Skip to main content
Close
Search
Help
Topics
Features
Blogs
Video
Podcasts
Magazine
Archive
Subscribe
Newsletter
About Us
Advertise
Login
Register
Facebook
YouTube
LinkedIn
Bluesky
American Scientist
Open navigation
Search
Help
Login
Tags
Astronautics
Aeronautics
Aesthetics
America250
Anatomy
Animal Behavior
Anthropogeography
Archaeology
Astrogeology
Astronautics
Astrophysics
Botany
Cartography
Chronology
Climatology
Cosmology
Crystallography
Cybernetics
Cytology
Ecology
Embryology
Epidemiology
Excerpt
Forestry
Geneaology
Genetics
Geographical Distribution
Geography
Geology
History Of Civilization
Human Ecology
Immunology
Information resources (general)
Information Theory
Linguistics
Logic
Metallurgy
Methodology
Microbiology
Mineralogy
Morphology
Motor Vehicles
Natural History
Nature Conservation
Neuroscience
Oceanography
Paleontology
Petrology
Philosophy of Science
Photography
Physiology
Poetry
Rationalism
Religion
Reproduction
Review
Scientists' Nightstand
Social Science
Special Collections
Statistics
Theology
Transportation
Virology
Zoology
Whose Cosmos Is It, Anyway?
Keivan G. Stassun
Astronomy
History of STEM
Sociology
Astronautics
Review
Scientists Nightstand
Books
Journey to the Stars
Jaime Herndon
History of STEM
Aeronautics
Astronautics
Review
Science Culture
Preparing for Apollo
Amy E. Foster
Astronomy
Astronautics
Scientists Nightstand
Books
The Costs of Being Sally Ride
Jaime Herndon
Physics
Astronautics
Review
Science Culture
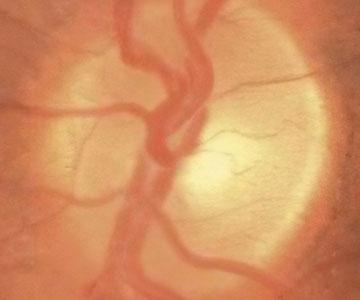
An Eye on Long-Term Space Flight
Shannon Persaud
Anthropology
Astronomy
Astronautics
Astrophysics
From The Staff
Do You Know Where Your Satellite Is Tonight?
David Finkleman
Astronomy
Communications
Engineering
Physics
Technology
Aeronautics
Astronautics
Astrophysics
Geography
Load more
×
AMSCI ICON NAVIGATION:
Navigation Menu
Search
Help
Log In, Register
My AmSci
Select Options
(not present on all pages)
Click "American Scientist" to access home page
×