Heart Waves
By Robert Frederick
Using artificial intelligence, researchers predict the propagation of the heart’s electrical signals.
Using artificial intelligence, researchers predict the propagation of the heart’s electrical signals.

DOI: 10.1511/2019.107.1.6
Sending a single shock to the heart to restore its regular rhythm is one of the most common procedures in modern medicine, and is used in emergency, non-emergency, and elective situations. But that shock, with as many as 360 joules of energy (the maximum setting of most external defibrillators), comes with risks, including damage to the heart. So for more than a decade researchers have been investigating the safety and effectiveness of a kind of low-energy defibrillation that involves a sequence of much weaker pulses.
“To choose, for example, the right frequency of these pulses or the right moment to apply them, this is a difficult task,” says Ulrich Parlitz of the Research Group Biomedical Physics at the Max Planck Institute for Dynamics and Self-Organization in Germany. Headed by Stefan Luther, this team of researchers has been seeking more insight into the mechanisms underlying the heart’s electrical dynamics, which can be chaotic during cardiac arrhythmia. They turned to mathematical models describing the heart’s electrical dynamics and a type of artificial intelligence called machine learning in which a computer (or, more properly, a “neural network” on a computer) progressively improves its performance by comparing its computed, predicted output to the ideal output (computed directly from the mathematical model). In a sense, then, the neural network learns, or is trained, without being explicitly programmed.
With his colleague Roland Zimmermann, Parlitz tried out a machine-learning approach with two relatively simple mathematical models describing the heart’s electrical dynamics. The first, simpler model has two variables. “So you train [the neural network] so that it reproduces these dynamics” with data for both variables, says Parlitz, and then test it by seeing how well it predicts the value of the second variable by providing it with data only from the first variable. In other words, says Parlitz, “Once it is trained, it provides you additional information about your underlying system.” That additional information is the sought-after insight into the heart’s electrical system.
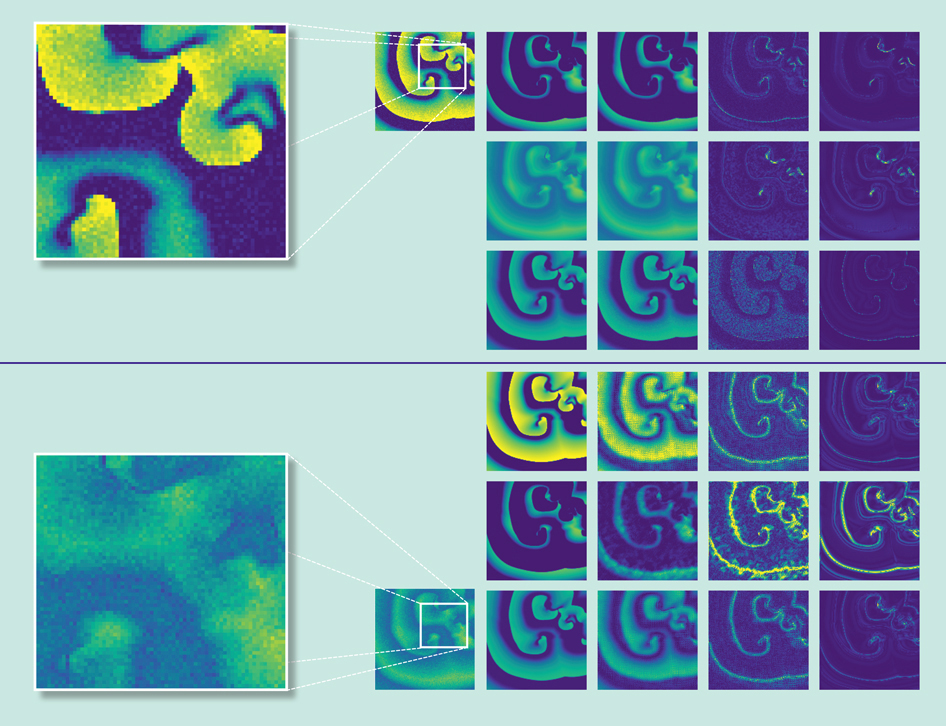
Adapted by Barbara Aulicino from data from R. S. Zimmerman and U. Parlitz..
In a real-life scenario, a medical professional may have easy access to only one data source, such as a patient’s electrocardiogram. By inputting the easily measured data, the idea behind the research is that a trained neural network could help medical professionals better predict heart conditions that are not so easily measured, such as abnormal flow of calcium currents inside the heart that can sometimes generate arrhythmias.
After proof-of-principle success with the simpler model, Zimmermann and Parlitz then trained their neural network with a more complicated mathematical model that uses four variables, testing how well it predicted three variables based on input from only one variable. They also introduced noise into their input data, which not only was a check of the robustness of the neural network’s predictions, but were also a kind of simulation of real-life data, which are always noisy. The results of two tests are summarized visually (above) and published in the April 24, 2018, issue of the journal Chaos. Parlitz says he is nearing publication of a different type of machine-learning algorithm—deep learning—that provides even higher resolution, but takes weeks, not hours, to train.
“The deep learning approach is extremely slow in learning,” says Parlitz, “but in the end you get better results” or further insights into the human heart via artificial intelligence.

Video courtesy of Roland Zimmermann and Ulrich Parlitz of the Research Group Biomedical Physics at the Max Planck Institute for Dynamics and Self-Organization.
An audio interview with Ulrich Parlitz:
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.