Science After the Volcano Blew
By Douglas Larson
Research near Mount St. Helens proceeded despite bureaucratic hurdles, limited funding and an extremely hazardous environment
Research near Mount St. Helens proceeded despite bureaucratic hurdles, limited funding and an extremely hazardous environment

DOI: 10.1511/2010.85.324
On May 20, 1980, two days after the catastrophic eruption of Mount St. Helens, U.S. Geological Survey (USGS) scientists landed in a helicopter near the volcano’s base, a short distance from what should have been the south shore of Spirit Lake. The lake, just 8 kilometers north-northeast of the volcano, was the largest of dozens of subalpine lakes battered by the eruption.

Photograph by the U.S. Forest Service.
Unable to see much that resembled a lake, the scientists suspected that Spirit Lake had either boiled away or was buried by avalanche debris. Further investigation revealed that the lake had survived, but it looked more like land than water because thousands of logs, tons of volcanic ash and other rubble blanketed its surface.
Volcanic activity and mudflows from Mount St. Helens created Spirit Lake roughly 3,000 years ago. But the 1980 eruption nearly blasted it to extinction. A debris avalanche, triggered by the eruption, slammed into the lake like an enormous tidal wave, sweeping water out of the lake basin northward and up the steep slopes of nearby Mount Margaret. Some water may have spilled over Margaret’s summit—about 800 meters above the lake’s surface. Most of the lake water rushed back down, stripping slopes of trees, rock and soil—down to bedrock—and washing all that was dislodged into the lake. Debris pushed the lake’s surface elevation 60 meters higher than its pre-eruption level of 975 meters, greatly altering its morphometry and blocking its natural outlet, the North Fork Toutle River. Previously kept in hydrological balance by that outlet, the lake had become locked in a hydrologically unstable basin by a debris dam 150 to 180 meters thick.

Photograph by the U.S. Forest Service.
Within hours of the eruption, pyroclastic flows, mudflows, heavy tephra fall and high-temperature geothermal waters poured into Spirit Lake, altering its waters. An initial survey described the lake as hypereutrophic, or oversupplied with nutrients; thoroughly anoxic; blackish in appearance; highly concentrated with metals and organic compounds; and containing extraordinarily prolific bacteria that reached densities thought to be unprecedented in natural aquatic systems. Phytoplankton, zooplankton and macroinvertebrates were almost nonexistent, with most taxa thought to be eliminated. Lacking oxygen, fish and other aquatic vertebrates perished. Considering these conditions, scientists predicted that the lake would take 20 years to recover, perhaps longer.
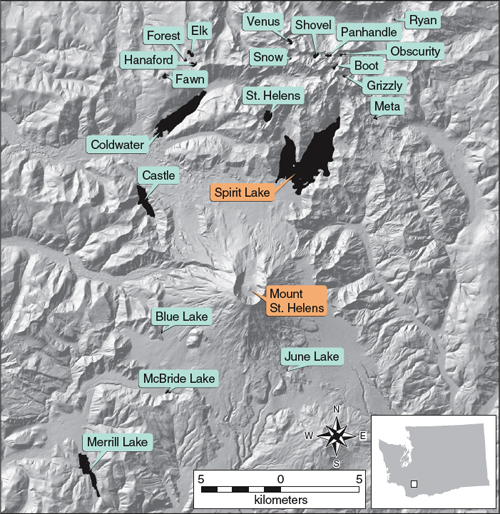
Map courtesy of Dianne McDonnell and James Thibault of the University of New Mexico.
I have spent the past 30 years observing the post-eruption response and recovery of Spirit Lake, first as a limnologist for the Portland District of the U.S. Army Corps of Engineers, and later, occasionally, as an independent researcher. Despite early predictions, the lake had made a remarkable recovery by 1986, which I reported in this magazine (see “The Recovery of Spirit Lake,” American Scientist, March–April 1993). In that article, I alluded to the unusual field-science experiences and challenges involved with working in a devastated landscape near the base of a still-active volcano. Over the past three decades, I have come to better appreciate the scale of those challenges and additional, human-made obstacles. Both took a toll on a once-in-a-lifetime scientific opportunity to study the microbial and chemical transformations—as well as the restoration—of a lake impacted so severely by a volcano. Needed research was accomplished. But more could and should have occurred. My hope is that by sharing a deeper explanation of my experiences, others will learn from the failures and the successes at Spirit Lake.
Various factors contributed to what became a limited effort to assess Spirit Lake: chiefly bureaucratic hindrance and inertia, a lack of funding, and unreliable logistical support in an incredibly challenging location. At the beginning, scientific efforts were spotty. The initial research on the eruption’s effects on Spirit Lake and other blast-zone lakes was suspended only four months after the eruption. Studies didn’t resume for nearly two years, when an emergency required them. Starting in late October 1980, with the onset of winter, Spirit Lake began rapidly filling with water. By April 1981, lake volume had increased by almost 30 percent, and its surface elevation had risen to roughly 1,017 meters. According to estimates by the Corps and the USGS, there was a high probability that the debris dam blocking the lake’s outlet would fail if lake levels rose above 1,055 meters. Failure of the dam would cause catastrophic flooding downstream, an event that would seriously threaten tens of thousands of people.

In November 1982, as a temporary solution, the Corps started pumping water from the lake to stabilize its surface elevation. Barge-mounted pumps removed lake water at the rate of 307 cubic meters per minute, discharging it across the debris dam through a pipeline 1,100 meters long that emptied into the North Fork Toutle River. Pumping during the first year held the lake level at just about the crucial 1,055 meters. Had the lake not been pumped, its surface elevation would have reached 1,079 meters by August 1983, causing the lake to overtop the debris dam or breach it at a lower elevation. Meanwhile, as a permanent solution, the Corps began constructing a tunnel outlet connecting Spirit Lake to the North Fork Toutle River via South Coldwater Creek.
But before any water could be released, the Corps was required to monitor the lake’s water quality. Post-eruption water-quality assessments had made it clear that Spirit Lake’s waters contained unusual biological and chemical pollutants. Releasing great amounts could contaminate public water supplies, disrupt fish that migrate from the ocean to freshwater to spawn, and imperil other valuable resources downstream. The U.S. Environmental Protection Agency, the U.S. Forest Service and the U.S. Public Health Service recommended that the Corps develop and maintain a long-term water- quality program at Spirit Lake to monitor the discharges.
In January 1983, three months after pumping commenced, I was assigned the task of developing and implementing the Corps’ Spirit Lake water- quality assessment. I was instructed to produce a “basic” program, one in which discharged water was simply collected periodically for bacteriological and chemical analyses. I argued that in order to comply with multiple federal directives, the Corps had to conduct wider limnological studies to systematically define the lake’s physical, chemical and biological properties for the purpose of tracking lake recovery, and to identify potentially hazardous chemical and biological constituents.
Yielding to these directives, the Corps finally approved the program, which was launched in April 1983. By then, unfortunately, lake conditions had already shifted substantially toward recovery, meaning that major post-eruption chemical and biological transformations had occurred without being thoroughly documented. Prior to 1983, Spirit Lake had been visited approximately 20 times for limnological data. But because of paltry logistical and funding support, these surveys were brief, lasting only a few hours and produced limited information. All visits but two were made in the summer and provided little data about seasonal differences in lake conditions.
Before more comprehensive research could begin at Spirit Lake, considerable planning was required. The eruption had created an immensely challenging set of geophysical obstacles, health hazards and dangerous lake conditions, all of which made for a unique limnological experience. The research required unorthodox logistics and sampling techniques that were sometimes more experimental than the lake science itself. Lake visits were needed year-round, with winter month surveys staged in deep snow, strong winds and icy waters.
During the early phase of lake research, 1980 through 1982, researchers used small rubber dinghies to collect water samples and deploy various recording instruments in situ. These fragile craft proved to be inadequate, for several reasons: They were too small to safely accommodate personnel and equipment; they lacked a cabin, exposing people to rain, wind, sun and blowing ash; they were unstable and difficult to control, especially in rough waters; and they could easily be pushed—even crushed—by the massive log rafts that drifted unpredictably across the lake.

Douglas W. Larson
In early 1983, I argued for a boat sturdy enough to resist collisions with floating logs and with sufficient engine power to avoid the logs or push them out of the way. Instead, I was furnished with a much-used fiberglass cabin cruiser (5.8 meters long) equipped with a castoff outboard motor. The only thing missing was a set of water skis. During a lake survey in May 1983, the boat struck a partially submerged log that produced a two-meter-long crack in the hull, causing the boat to slowly fill with water. While three fellow limnologists bailed frantically, I managed to steer the boat to shore before it could sink. The boat was deemed “unserviceable” and trucked out for disposal.
The near sinking of the cabin cruiser in frigid waters prompted me to file a complaint with the Corps. The Corps authorized the use of its river-survey boat, christened Traveler, for lake research at Mount St. Helens, along with the purchase of survival suits to be worn in case someone fell overboard or the boat sank. Wearing the suits gave me and Patrick Cagney, my frequent field assistant, a sense of security, if only a false one: Had we somehow ended up in the lake, bobbing around in frigid water, the suits would have delayed hypothermia, but floating logs would likely have crushed us.

Hailed as a steel-hulled vessel, the 8-meter-long Traveler was airlifted into Spirit Lake by a U.S. Army twin-rotor Chinook helicopter based at Fort Lewis, Washington. The underside of the boat’s hull had been retrofitted with a cage-like apparatus that enclosed the rudder and propeller assembly. On paper anyway, this jerry-built device was supposed to protect the boat from being immobilized by floating logs and other debris hazards. Nevertheless, despite our cautious operation procedures, collisions with submerged logs were inevitable. Moreover, unforeseeable and sudden shifts in wind direction and velocity often drove logs en masse toward the boat while it was anchored at a lake sampling station. Most of these wind-propelled logs bypassed the boat, but some struck the hull. Frequently, the boat was rammed through log rafts for distances ranging from a few hundred meters to a kilometer or two to reach sampling stations or to return to the boat’s moorage at day’s end.
We made a disturbing discovery in October 1983: Despite its acclaimed steel hull, Traveler was leaking. In the time between field trips to Spirit Lake, usually a period of one to two weeks, the leaks were sufficient to produce a half-meter-deep pool of water in the bottom of the hull. Weighted down by this water, Traveler settled into mud at its moorage. Raising the craft required those aboard to use the boat’s bilge pumps for at least 30 minutes before getting underway.
As with all of our endeavors at Spirit Lake, hope sprang eternal that we could carry on despite such challenges. Optimism was not enough, however. During a field trip on October 28, 1983, the leaks—combined with bureaucratic red tape and copious paperwork—nearly led to a disaster. Shoving off that day, I was accompanied by two upper-management engineers from the Corps’ North Pacific Division in Portland. Aboard to inspect my lake-research program, both men were looking forward to a pleasant day on the lake.
Shortly after reaching a sampling station around 10 a.m., the wind shifted and began to push the log raft in our direction. Soon, the boat was surrounded by logs and was taking on water. I activated the bilge pumps, but they worked for only a short time. One of my passengers discovered that the batteries that powered the pumps were dead. Hemmed in by logs, with the water filling the boat’s hull and no life raft, we found ourselves trapped on a sinking craft.
Using the boat’s radio, I contacted a Corps employee at the Spirit Lake pumping station and informed him of our predicament. I asked if he could send his helicopter to Longview, Washington, a half-hour flight away, to obtain new batteries. He replied yes but said he would first need permission from his supervisor in Portland. I told him to hurry; Our boat was sinking. About an hour later, he radioed back to say he had permission to send the helicopter but that I would need a typed purchase order signed by my immediate supervisor in Portland. Recalling the glacial speed that purchase orders usually progressed through the bureaucracy, I replied that the urgency of our situation called for a more expedient process. Finally the purchasing department authorized an emergency purchase order that was telephoned to a battery dealer in Longview. The batteries reached us around 4 p.m.
Two weeks later, again while encircled by logs, Traveler began to take on more water than usual, causing the boat to settle by the stern. Lake depth at this location was about 30 meters, sufficiently deep to make salvage extremely difficult, if not impossible. Lake-water temperature was 8 degrees Celsius, cold enough to induce hypothermia had I been forced to abandon ship. The bilge pumps evacuated some of the water bubbling up from the engine room’s deck, but water seemed to be coming in faster than ever before. I radioed the helicopter and informed the pilot of the problem. He immediately flew to my position and hovered nearby, waiting to rescue me if the boat capsized or sank. Reassured by my overhead escort, I broke through a kilometer-wide raft of logs before reaching open water. The rest of the journey became a race to reach the moorage before Traveler sank. With Traveler listing to one side, and the throttle wide open, the boat plowed into the lake bottom a few meters from shore. I waded to dry land.
The protective cage was no match for Spirit Lake. An inspection revealed that water had entered the boat through holes that had been drilled in the hull to attach the cage. Repeated blows from submerged and floating logs had loosened the cage’s bolts, breaking their watertight seals. In April 1984, Traveler was unceremoniously hauled back to Portland and retired from service, its “indestructible” designation left in doubt.
Then there was the matter of shelter. For four years after the eruption, we conducted laboratory tasks—sample preparations, packaging and analyses—out in the open. We were exposed to blowing ash and searing heat in summer, drifting snow and bitter cold in winter and gale-force winds year-round. Helicopters whipped up great clouds of ash, which enveloped us as we huddled around delicate laboratory equipment, trying to keep precious water samples from being contaminated.

Douglas W. Larson
After I reported that these conditions posed a health hazard and compromised scientific precision and accuracy, the Corps authorized the construction of a field laboratory at Spirit Lake. A Sikorsky helicopter airlifted a small fiberglass Quonset hut to the south shore of the lake on June 14, 1984. About 4 by 2.5 meters, the building was equipped with storage shelves on one side and a laboratory bench on the other. Outside, we reinforced it with a 1.5-meter-high earthen berm, which we enclosed with a wall of roughly 600 sandbags. (Throughout the lab’s existence, we filled and stacked sandbags. In time, I began asking all visitors to fill 10 sandbags each before boarding a helicopter to begin their trip away from Spirit Lake. I’d say: “No sandbags, no boarding pass.” No one refused.)

Douglas W. Larson
The shelter greatly increased our safety and comfort, especially during winter when we could retreat to the lab’s warm interior to avoid hypothermia. Also, boat gear, outboard motors, laboratory equipment, analytical supplies, propane stoves, survival suits, parachute flares, emergency rations and other essential materials could be left at the site, thus largely removing weight as a constraint on helicopter flights. As required by the U.S. Forest Service, which managed the Mount St. Helens National Volcanic Monument, the lab was painted with camouflage to be “inconspicuous.” On that terrain, however, the laboratory was anything but. It was the only bit of color in an unrelieved expanse of vast grey mudflow and bleached, broken skeletons of tree trunks strewn across the lakeshore.
Helicopter transportation was essential to scientific research on blast-zone lakes and rivers, including Spirit Lake. The helicopters provided rapid, safe transport to and from scattered study sites, some of which were inaccessible by road vehicles. Many of the pilots had learned to fly while serving in the military during the Vietnam War and could execute unconventional operations that required innovation and daring. Milt Walker, a former U.S. Army helicopter pilot, once flew us to Spirit Lake during a snowstorm, then returned several hours later in near-zero visibility to take us home. Another veteran pilot regularly landed his jumbo Sikorsky helicopter on an island of loose avalanche debris in Spirit Lake, waiting for us to disembark while his craft perched precariously on the island’s summit.

In addition, helicopters provided a quick and reliable means of escape if someone was hurt or if the volcano stirred. The importance of having a means of evacuation was made clear during the summer of 1983. While en route to a Spirit Lake sampling station aboard Traveler, a USGS scientist who was a member of our research team tried to open a reagent bottle containing a mixed solution of highly concentrated sodium hydroxide, sodium iodide and sodium azide. The bottle’s cap suddenly blew off, spraying her face with caustic reagents. Blinded and screaming, she fell to the deck. Frantic, we scooped water from the lake and splashed it across her face, trying to flush the burning chemicals from her eyes and skin. I radioed a medical evacuation request to our helicopter pilot, who was heading to the airport at Longview, Washington, to refuel. When we reached the boat moorage, the helicopter was waiting with its rotor blades spinning, ready for immediate takeoff. The injured researcher and a fellow USGS scientist were flown to a hospital in Longview, about 65 air-kilometers away. Within minutes, she was in the hospital’s emergency room undergoing treatment. She fully recovered from her burns.

Douglas W. Larson
In June 1984, we tried another approach to getting where we needed to be on the lake to collect our samples. I obtained a 5-meter-long Zodiac inflatable boat, which was much sturdier than the dinghies used in the initial Spirit Lakes studies. The new boat was also equipped with a harness that allowed a helicopter to lift it over the log raft into the lake’s open-water area. Strong east winds regularly pushed the log raft against the shore where our field lab was located, making it impossible to launch our boat. After the boat was lowered into the lake and anchored, the helicopter returned to the lab and delivered us to the waiting boat. If, at day’s end, the logs had not moved, the helicopter flew out to our position and lifted us back to the lab before retrieving the boat.

Patrick Cagney
When the wind was blowing strongly and unpredictably, the log raft would often surround us and press in from all sides, threatening to crush both boat and crew. On one occasion, as we sat trapped amidst logs scraping ominously on the boat’s rubber hull, I radioed our pilot and urgently requested that he extract us. Demonstrating considerable skill, he set one of the chopper’s skids on the edge of the boat and held his craft steady as my two colleagues climbed aboard. Revving up the engine for takeoff, the pilot waved goodbye and instantly rose skyward, blasting me with a brief but tornado-like downburst of air. The helicopter returned within minutes, and while it hovered overhead, the pilot slowly deployed a cable, which I grabbed when it got within reach and attached to the boat’s harness. He then dropped the helicopter close to the boat and motioned to me to climb aboard. I pulled myself into the helicopter and crouched in the door, looking down at the boat and directing the pilot as we ascended straight up. I signaled to him when the cable was taut. The boat rose suddenly from the log raft and swung beneath us as we returned to shore.
Not all of the hazards encountered in the blast zone were readily perceived. The black, toxic waters of lakes and rivers harbored potentially pathogenic bacteria including Klebsiella pneumoniae, Pseudomonas aeruginosa, and Legionella pneumophila. L. pneumophila had caused the mysterious pneumonia epidemic at an American Legion convention in Philadelphia in July 1976. Called Legionnaires’ disease, the illness sickened 211 people, 34 of whom died.

Patrick Cagney
In April 1981, the State of Washington’s Department of Social and Health Services (WDSHS) was notified that eight scientists had become ill with fever, cough and sore throat after working around the lakes and rivers in the blast zone. Further inquiry found that among 250 scientists who had visited the blast zone between June 1980 and June 1981, 28 reported developing headaches, sore throat and high fever (103–104 degrees Fahrenheit) within a week after visiting the blast zone. All recovered within a few days.
Field scientists suggested that the puzzling illness was Legionnaires’ disease. John Kline of the USGS reported that several members of his agency’s research teams had gotten sick with flulike symptoms soon after visiting lakes and rivers in the blast zone. He indicated that water samples collected from Spirit Lake and the North Fork Toutle River contained “extremely high concentrations” of Legionella bacteria, with one sample yielding 1.3 × 108 organisms per liter. Spirit Lake and other blast-zone waters provided an ideal medium for explosive bacterial growth, most likely due to nutrient enrichment, increased water temperatures from geothermal springs and other factors such as the waters’ exposure to volcanic debris.
Subsequent research by David Tison of Oregon State University and colleagues lent credence to allegations that the mysterious illness was Legionnaires’ disease. They found relatively large numbers (up to 1.3 × 107 organisms per liter) of potentially virulent L. pneumophila in Spirit Lake and other blast zone waters during the spring and summer of 1981. Michael Glass, a WDSHS microbiologist, reported that L. pneumophila was present in 52 percent of samples—numbering in the hundreds—collected from Spirit Lake between 1983 and 1986. Glass identified six other Legionella species, two of which (L.gormanii and L. longbeachae) are also human pathogens. Two new species were named: Legionella sainthelensi and L. spiritensis.

Douglas W. Larson
Public health investigators noted that people who contracted the illness were mostly scientists working in close proximity to blast-zone lakes and rivers. They hypothesized that mists generated at these sites by turbulent streams and waterfalls and, especially, the rotor blasts of nearby helicopters enabled exposure to the disease organisms. Legionnaires’ disease is not caused merely by the presence of Legionella, but by contact with bacteria-bearing aerosols in mist-filled air that people breath in.
The occurrence of Legionnaires’ disease was never proven, although the WDSHS conceded that the illness could have been a “mild form” of the disease, which the agency called “Red Zone Fever.” Had Legionnaires’ disease been confirmed, state and federal health authorities would have closed the blast zone for months or longer until the disease threat had abated. Closure would have put the Corps at loggerheads with its Congressional mandate in the blast zone—that is, to control flooding and mudflows, remove sediment blocking navigable rivers, repair or replace bridges and roads and provide damage assessments, all without delay to protect lives and property. Ironically, the threat of Legionnaires’ disease was a major hook that got us more money for lake research.
By 1986, as funding ended, our work at Spirit Lake was winding down. I estimate that the Corps spent about $2 million on lake research in the Mount St. Helens blast zone between 1980 and 1986. By then, my colleagues and I had collected data there about 100 times, measuring temperature and other physical properties; analyzing water chemistry; and determining species composition and relative abundance of phytoplankton, zooplankton and bacteria. On September 29, 1986, the field lab was stripped of its contents and airlifted out. After that, funding for limnological research for Spirit Lake was virtually unavailable, even though the lake is a key feature of the Mount St. Helens National Volcanic Monument. Consequently, only about a dozen trips were made to collect limnological data at the lake between 1986 and 2005—all of them, as far as I know, by me and Richard Petersen of Portland State University.

None of this later work amounted to long-term, coherent, methodical research, as it should have. Spirit Lake has a unique status as a large body of water that was severely transformed by a massive volcanic eruption and that was also the source of discharge into another waterway. If we had obtained the full funding the project deserved, we could have broadened our limnological scope by studying nutrient recycling and water optics in greater detail. We also could have observed the ways that physical, chemical and biological factors influenced rates of photosynthesis in the lake as well as the distribution of energy though the aquatic food chain.
Recently the ecologist and conservation biologist David Lindenmayer of the Australian National University and others argued that it is necessary to better prepare for scientific study of effects from catastrophic ecological disturbances before they happen. The fact that Spirit Lake was barely studied during the first three years after the 1980 eruption and then again after 1986 is a case in point that better planning is needed. Lindenmayer and colleagues, in the journal Frontiers in Ecology and the Environment, proposed that adequate funding, operational plans and infrastructure (logistics and equipment) be made available in advance of such a disturbance.
This argument has merit, of course, but my experience at Spirit Lake after the 1980 eruption taught me that other factors can hinder research after a natural disaster: Bureaucratic foot-dragging, interagency rivalries and poor judgment can forestall essential research. Funding, planning and infrastructure may be available in advance, but how these elements are deployed is another question. Although Congress appropriated millions, if not billions, of dollars for projects in the vicinity of Mount St. Helens, only a tiny fraction ended up financing research.
The challenges we encountered in the blast zone were often simply unpredictable, requiring on-the-spot innovation. Most of the effort I made at Spirit Lake was devoted to solving unforeseen logistical and operational problems that stood in the way of the science. No amount of planning would have prepared me or my colleagues for these contingencies. Problem solving was accomplished largely through trial and error. Our efforts to study Spirit Lake were propelled ultimately by reasoning, initiative, creativity, enthusiasm and, perhaps most important, an abiding belief that it could be done. Any scientist determined to conduct research in a natural disaster zone would be well advised to pack the same qualities before departing for the field.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.