Gauss's Day of Reckoning
By Brian Hayes
A famous story about the boy wonder of mathematics has taken on a life of its own
A famous story about the boy wonder of mathematics has taken on a life of its own

DOI: 10.1511/2006.59.200
Let me tell you a story, although it's such a well-worn nugget of mathematical lore that you've probably heard it already:
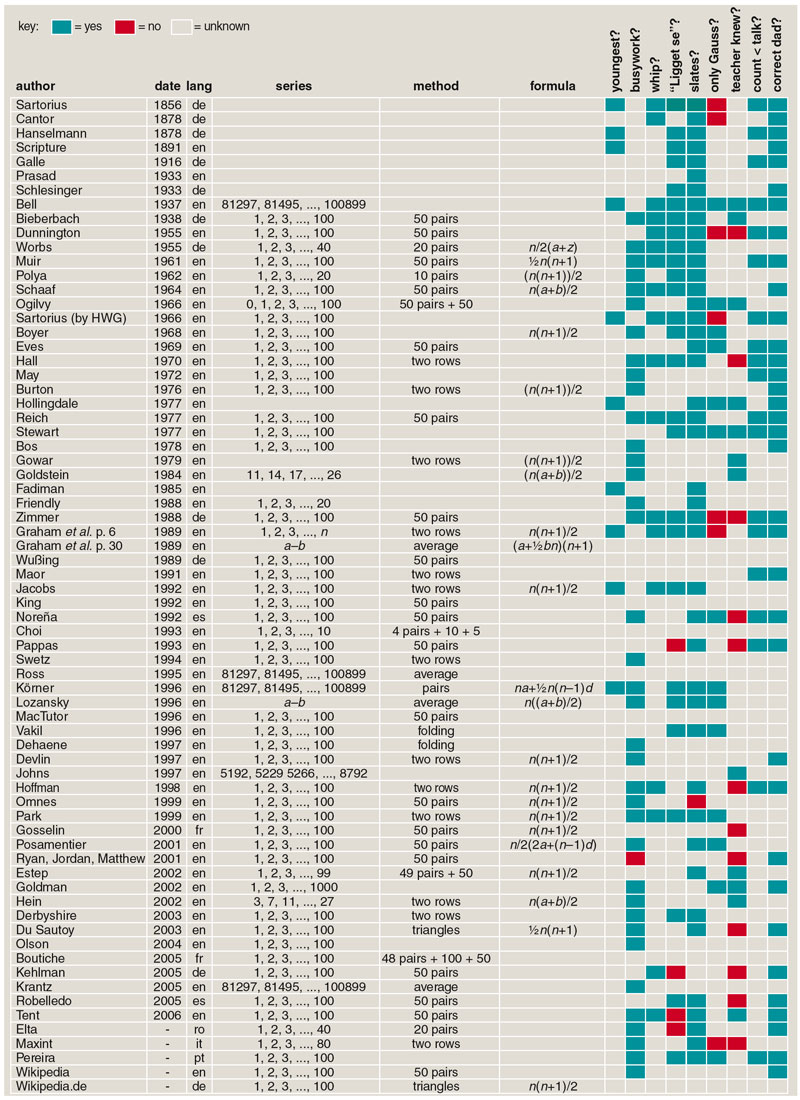
In the 1780s a provincial German schoolmaster gave his class the tedious assignment of summing the first 100 integers. The teacher's aim was to keep the kids quiet for half an hour, but one young pupil almost immediately produced an answer: 1 + 2 + 3 + ... + 98 + 99 + 100 = 5,050. The smart aleck was Carl Friedrich Gauss, who would go on to join the short list of candidates for greatest mathematician ever. Gauss was not a calculating prodigy who added up all those numbers in his head. He had a deeper insight: If you "fold" the series of numbers in the middle and add them in pairs—1 + 100, 2 + 99, 3 + 98, and so on—all the pairs sum to 101. There are 50 such pairs, and so the grand total is simply 50×101. The more general formula, for a list of consecutive numbers from 1 through n, is n(n + 1)/2.
The paragraph above is my own rendition of this anecdote, written a few months ago for another project. I say it's my own, and yet I make no claim of originality. The same tale has been told in much the same way by hundreds of others before me. I've been hearing about Gauss's schoolboy triumph since I was a schoolboy myself.

Illustration by Theoni Pappas, reprinted from Pappas 1993 by permission.
The story was familiar, but until I wrote it out in my own words, I had never thought carefully about the events in that long-ago classroom. Now doubts and questions began to nag at me. For example: How did the teacher verify that Gauss's answer was correct? If the schoolmaster already knew the formula for summing an arithmetic series, that would somewhat diminish the drama of the moment. If the teacher didn't know, wouldn't he be spending his interlude of peace and quiet doing the same mindless exercise as his pupils?
There are other ways to answer this question, but there are other questions too, and soon I was wondering about the provenance and authenticity of the whole story. Where did it come from, and how was it handed down to us? Do scholars take this anecdote seriously as an event in the life of the mathematician? Or does it belong to the same genre as those stories about Newton and the apple or Archimedes in the bathtub, where literal truth is not the main issue? If we treat the episode as a myth or fable, then what is the moral of the story?
To satisfy my curiosity I began searching libraries and online resources for versions of the Gauss anecdote. By now I have over a hundred exemplars, in eight languages. (The collection of versions is available here.) The sources range from scholarly histories and biographies to textbooks and encyclopedias, and on through children's literature, Web sites, lesson plans, student papers, Usenet newsgroup postings and even a novel. All of the retellings describe what is recognizably the same incident—indeed, I believe they all derive ultimately from a single source—and yet they also exhibit marvelous diversity and creativity, as authors have struggled to fill in gaps, explain motivations and construct a coherent narrative. (I soon realized that I had done a bit of ad lib embroidery myself.)
After reading all those variations on the story, I still can't answer the fundamental factual question, "Did it really happen that way?" I have nothing new to add to our knowledge of Gauss. But I think I have learned something about the evolution and transmission of such stories, and about their place in the culture of science and mathematics. Finally, I also have some thoughts about how the rest of the kids in the class might have approached their task. This is a subject that's not much discussed in the literature, but for those of us whose talents fall short of Gaussian genius, it may be the most pertinent issue.
I started my survey with five modern biographies of Gauss: books by G. Waldo Dunnington (1955), Tord Hall (1970), Karin Reich (1977), W. K. Bühler (1981) and a just-issued biography by M. B. W. Tent (2006). The schoolroom incident is related by all of these authors except Bühler. The versions differ in a few details, such as Gauss's age, but they agree on the major points. They all mention the summation of the same series, namely the integers from 1 to 100, and they all describe Gauss's method in terms of forming pairs that sum to 101.
None of these writers express much skepticism about the anecdote (unless Bühler's silence can be interpreted as doubt). There is no extended discussion of the story's origin or the evidence supporting it. On the other hand, references in some of the biographies did lead me to the key document on which all subsequent accounts seem to depend.
This locus classicus of the Gauss schoolroom story is a memorial volume published in 1856, just a year after Gauss's death. The author was Wolfgang Sartorius, Baron von Waltershausen, professor of mineralogy and geology at the University of Göttingen, where Gauss spent his entire academic career. As befits a funerary tribute, it is affectionate and laudatory throughout.
In the portrait Sartorius gives us, Gauss was a wunderkind. He taught himself to read, and by age three he was correcting an error in his father's arithmetic. Here is the passage where Sartorius describes Gauss's early schooling in the town of Braunschweig, near Hanover. The translation, except for two phrases in brackets, is by Helen Worthington Gauss, a great-granddaughter of the mathematician.
In 1784 after his seventh birthday the little fellow entered the public school where elementary subjects were taught and which was then under a man named Büttner. It was a drab, low school-room with a worn, uneven floor.... Here among some hundred pupils Büttner went back and forth, in his hand the switch which was then accepted by everyone as the final argument of the teacher. As occasion warranted he used it. In this school—which seems to have followed very much the pattern of the Middle Ages—the young Gauss remained two years without special incident. By that time he had reached the arithmetic class in which most boys remained up to their fifteenth year.
Here occurred an incident which he often related in old age with amusement and relish. In this class the pupil who first finished his example in arithmetic was to place his slate in the middle of a large table. On top of this the second placed his slate and so on. The young Gauss had just entered the class when Büttner gave out for a problem [the summing of an arithmetic series]. The problem was barely stated before Gauss threw his slate on the table with the words (in the low Braunschweig dialect): "There it lies." While the other pupils continued [counting, multiplying and adding], Büttner, with conscious dignity, walked back and forth, occasionally throwing an ironical, pitying glance toward this the youngest of the pupils. The boy sat quietly with his task ended, as fully aware as he always was on finishing a task that the problem had been correctly solved and that there could be no other result.
At the end of the hour the slates were turned bottom up. That of the young Gauss with one solitary figure lay on top. When Büttner read out the answer, to the surprise of all present that of young Gauss was found to be correct, whereas many of the others were wrong.
Incidental details from this account reappear over and over in later tellings of the story. The ritual of piling up the slates is one such feature. (It must have been quite a teetering heap by the time the hundredth slate was added!) Büttner's switch (or cane, or whip) also made frequent appearances until the 1970s but is less common now; we have grown squeamish about mentioning such barbarities.
What's most remarkable about the Sartorius telling of the story is not what's there but what's absent. There is no mention of the numbers from 1 to 100, or any other specific arithmetic progression. And there is no hint of the trick or technique that Gauss invented to solve the problem; the idea of combining the numbers in pairs is not discussed, nor is the formula for summing a series. Perhaps Sartorius thought the procedure was so obvious it needed no explanation.
A word about the bracketed phrases: Strange to report, the Worthington Gauss translation does mention the first 100 integers. Where Sartorius writes simply "eine arithmetischen Reihe," Worthington Gauss inserts "a series of numbers from 1 to 100." I cannot account for this interpolation. I can only guess that Worthington Gauss, under the influence of later works that discuss the 1-to-100 example, was trying to help out Sartorius by filling in an omission. The second bracketed passage marks an elision in the translation: Where Sartorius has the pupils "rechnen, multiplizieren und addieren," Worthington Gauss writes just "adding." I'll have more to say on this point below.
If Sartorius did not specify a series running from 1 to 100, where did those numbers come from? Could there be some other document from Gauss's era that supplies the missing details? Perhaps someone to whom Gauss told the story "with amusement and relish" left a record of the occasion. The existence of such a corroborating document cannot be ruled out, but at present there is no evidence for it. None of the works I have seen makes any allusion to another early source. If an account from Gauss's lifetime exists, it remains so obscure that it can't have had much influence on other tellers of the tale.
In the literature I have surveyed, the 1-100 series makes its first appearance in 1938, some 80 years after Sartorius wrote his memoir. The 1-100 example is introduced in a biography of Gauss by Ludwig Bieberbach (a mathematician notorious as the principal instrument of Nazi anti-Semitism in the German mathematical community). Bieberbach's telling of the story is also the earliest I have seen to specify Gauss's strategy for calculating the sum—the method of forming pairs that add to 101. Should Bieberbach therefore be regarded as the source from whom scores of later authors have borrowed these "facts"? Or is this a case of multiple independent invention?
If you think it utterly implausible that two or more authors would come up with the same example and the same method, then Bieberbach himself is disqualified as the source. A full millennium before Gauss and Büttner had their classroom confrontation, essentially the same problem and solution appeared in an eighth-century manuscript attributed to Alcuin of York.
Brian Hayes
Furthermore, in the years since Bieberbach wrote, there is unmistakable evidence of independent invention. Not all versions agree that the sequence of numbers was the set of consecutive integers from 1 through 100. Although that series is the overwhelming favorite, many others have been proposed. Some are slight variations: 0-100 or 1-99. Several authors seem to feel that adding up 100 numbers is too big a job for primary-school students, and so they trim the scope of the assignment, suggesting 1-80, or 1-50, or 1-40, or 1-20, or 1-10. A few others apparently think that 1-100 is too easy, and so they give 1-1,000 or else a series in which the difference between successive terms is a constant other than 1, such as the sequence 3, 7, 11, 15, 19, 23, 27. (The example series chosen by various authors and other features of the versions are tabulated in the table above.)
Perhaps the most influential version of the story after that of Sartorius is the one told by Eric Temple Bell in Men of Mathematics, first published in 1937. Bell has a reputation as a highly inventive writer (a trait not always considered a virtue in a biographer or historian). He turns the Braunschweig schoolhouse into a scene of gothic horror: "a squalid relic of the Middle Ages run by a virile brute, one Büttner, whose idea of teaching the hundred or so boys in his charge was to thrash them into such a state of terrified stupidity that they forgot their own names." Very cinematic! When it comes to the arithmetic, however, Bell is one of the few writers who scruple to distinguish between fact and conjecture. He doesn't claim to know the actual numerical series, but writes: "The problem was of the following sort, 81297 + 81495 + 81693 + ... + 100899, where the step from one number to the next is the same all along (here 198), and a given number of terms (here 100) are to be added." (Personally, I'd have a hard time even writing that problem on a small slate, much less solving it.)
It's a challenge to sort out patterns of influence and transmission in such a collection of stories. When a later author mentions the series 81297 + 81495 + ..., we can be pretty sure those numbers came from Bell. When the example given is 1-100, however, it's not so easy to trace the line of inheritance—if there is one. And the dozen or so other sequences that appear in the literature argue for a high rate of mutation; every one of those examples had to be invented at least once.
Tellers of a tale like this one seem to work under a special dispensation from the usual rules of history-writing. Authors who would not dare to alter a fact such as Gauss's place of birth or details of his mathematical proofs don't hesitate to embellish this anecdote, just to make it a better story. They pick and choose from the materials available to them, taking what they need and leaving the rest—and if nothing at hand suits the purpose, then they invent! For example, several authors show a familiarity with Bell's version of the story, quoting or borrowing distinctive phrases from it, but they decline to go along with Bell's choice of a series beginning 81297, falling back instead on the old reliable 1-100 or inserting something else entirely. Thus it appears that what is driving the evolution of this story is not just the accumulation of errors of transmission, as in the children's game "whisper down the lane"; authors are deliberately choosing to "improve" the story, to make it a better narrative.
For the most part, I would not criticize this practice. Effective storytelling is surely a legitimate goal, and outside of formal scholarly works, a bit of embroidery on the bare fabric of the plot does no harm. A case in point is the theme of "busywork" found in most recent tellings of the story (including mine). It seems we feel a need to explain why Büttner would give his pupils such a long and dreary exercise. But Sartorius says nothing at all about Büttner's motivation, nor do any of the other 19th-century works I've consulted. The idea that he wanted to keep the kids quiet while he took a break is entirely a modern inference. It's probably wrong—at best it's unattested—and yet it answers a need of readers today.
In the same spirit, many authors confront the question that got me started on this quest: How did Büttner do the math? Bell is adamant that Büttner knew the formula beforehand; others say he learned the trick only when Gauss explained it to him. An example of the latter position is the following account written in 2001 by three fifth-grade students, Ryan, Jordan and Matthew:
When Gauss was in elementary school his teacher Master Büttner did not really like math so he did not spend a lot of time on the subject. One of the problems his teacher gave the class was "add all the whole numbers from 1 to 100". His teacher Master Büttner was amazed that Gauss could add all the whole numbers 1 to 100 in his head. Master Büttner didn't believe Gauss could do it, so he made him show the class how he did it. Gauss showed Master Büttner how to do it and Master Büttner was amazed at what Gauss just did.
Am I being unfair in matching up Eric Temple Bell against three fifth-graders? Unfair to which party? Both offer interpretations that can't be supported by historical evidence, but Ryan, Jordan and Matthew are closer to the experience of classroom life.
As with the identity of the series, the details of how Gauss solved the problem remain a matter of conjecture. The algorithm that I suggested—folding the sequence in half, then adding the first and last elements, the second and next-to-last, etc.—is not the only possibility. A related but subtly different algorithm is mentioned by many authors. The idea is to write down the series twice, once forward and once backward, and then add corresponding elements. For the familiar series 1-100 this procedure yields 100 pairs of 101, for a total of 10,100; then, since the original series was duplicated, we need to divide by 2, arriving at the correct answer 5,050. The advantage of this scheme is that it works the same whether the length of the sequence is odd or even, whereas the folding algorithm requires some fussy adjustments to deal with an odd-length series.
Brian Hayes
A third approach to the summation problem strikes me as better still. The root idea is that for any finite set of numbers, whether or not the numbers form an arithmetic progression, the sum is equal to the average of all the elements multiplied by the number of elements. Thus if you know the average, you can easily find the sum. For most sets of numbers, this fact is not very useful, because the only way to calculate the average is first to calculate the sum and then divide by the number of elements. For an arithmetic progression, however, there is a shortcut: The average over the entire series is equal to the average of the first and last elements (or the average of any other elements symmetrically arrayed around the midpoint). If this was Gauss's secret weapon, then his mental multiplication was not 50 x 101 but 100 x 50½.
All three of these ideas—and a few more besides—have been presented by one author or another as the method that Gauss discovered during his first arithmetic lesson. Expressed as formulas for summing consecutive integers from 1 through n, the three rules (folding, double rows, average) look like this:
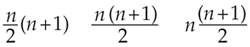
Mathematically, it's obvious they are equivalent: For the same value of n, they produce the same answer. But the computational details are different and, more important, so are the reasoning processes that lead to these formulas.
There is yet another way of thinking about the summation process: n(n + 1)/2 has been known since antiquity as the formula for the triangular numbers, those in the sequence 1, 3, 6, 10, 15, 21.... Thus some authors suggest that Gauss was thinking geometrically, forming an n-by-n + 1 rectangle and cutting it along the diagonal.
So much for how the prodigious Carl Friedrich Gauss solved the problem. What about the rest of the students in the class? Let me invite you to take a sheet of paper and actually try adding the numbers from 1 to 100.
Finished? Already?
What I discovered when I tried this experiment is that it's really hard to do it the hard way. You may set out to plod dutifully through all the addition operations, but shortcuts present themselves even when you're not looking for them. Suppose you adopt the standard primary-school algorithm, writing down all 100 numbers in a tall column and then starting work on the units digits. After the first 10 digits, the partial sum is 45; the next 10 digits add another increment of 45, bringing the partial sum to 90; then 45 more makes 135, and so on. How far would a student get in this process before recognizing a repetitive pattern? On turning to the tens digits, the pattern is even harder to miss: There are ten 1s followed by ten 2s, then ten 3s, etc. Surely any student who has the skills to complete this task at all would not add those repeated numbers one by one. A more likely strategy would be the one Sartorius implied when he wrote "count, multiply and add"—the phrase that Helen Worthington Gauss reduced to mere "adding."
On a small slate or a sheet of paper, it's difficult to write 100 numbers in a column, and so students would likely break the task down into subproblems. Suppose you start by adding the numbers from 1 to 10, for a sum of 55. Then the sum of 11 through 20 is 155, and 21 through 30 yields 255. Again, how far would you continue before spotting the trend?
Admittedly, these shortcuts can't match the elegance and ingenuity of Gauss's method. They are tied to the decimal representation of numbers, and they also don't generalize as well to arithmetic progressions other than lists of consecutive integers. But they do remind us that there's usually more than one good way to solve a problem.
I suspect that only one kind of student would ever be likely to add the numbers from 1 through 100 by performing 99 successive additions—namely, a student using a computer or a programmable calculator. And for that student, the simplest strategy might in fact be the best one.
We can hope that a modern Büttner—deprived of his whip, of course, and teaching in a classroom where computers have replaced slates—would not be drilling students on skills of such dubious utility as adding up a long series of numbers by hand. But the new Büttner just might ask his pupils to write a program to calculate the sum of any arithmetic progression. A new Gauss, with the same keen insight, could create a very efficient program based on the pairing idea—and that feat still deserves the highest admiration. But the modern Gauss might not be the first to fling his or her laptop on the table and cry "There it lies!" Writing that clever program—and testing and debugging it, and proving its correctness—would be no quicker than writing the straightforward step-by-step version. In this respect, technology may be something of an equalizer.
The story of Gauss and his conquest of the arithmetic series has a natural appeal to young people. After all, the hero is a child—a child who outwits a "virile brute." For many students, that is surely an inspiration. But I worry a little that the constant repetition of stories like this one may leave the impression that mathematics is a game suited only to those who go through life continually throwing off sparks of brilliance.
On first hearing this fable, most students surely want to imagine themselves in the role of Gauss. Sooner or later, however, most of us discover we are one of the less-distinguished classmates; if we eventually get the right answer, it's by hard work rather than native genius. I would hope that the story could be told in a way that encourages those students to keep going. And perhaps it can be balanced by other stories showing there's a place in mathematics for more than one kind of mind.
In collecting versions of the Gauss anecdote I've been helped by dozens of librarians as well as friends and others. I particularly want to thank Johannes Berg of the University of Cologne; Caroline Grey of the Johns Hopkins University libraries; Stephan Mertens of the University of Magdeburg; Ivo Schneider of the Bundeswehr University, Munich; Margaret Tent of the Altamont School in Birmingham, Alabama, and Mary Linn Wernet of the Northwestern State University libraries in Natchitoches, Louisiana.
© Brian Hayes
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.