The Biodiversity Conservation Paradox
By Mark Vellend
Even in places where nature is perceptibly altered by human actions, the number of species does not necessarily decline.
Even in places where nature is perceptibly altered by human actions, the number of species does not necessarily decline.

Since the arrival of Homo sapiens in New Zealand sometime during the 13th century, the number of plant species in the country has doubled, from roughly 2,000 to more than 4,000. This island nation with no native land mammals apart from a few species of bats is now home to more than two dozen types of mammals, including possums, rabbits, deer, and wallabies. Biodiversity has increased. And yet New Zealand, along with the rest of the Earth, is often described as undergoing a “biodiversity crisis,” with species going extinct at an alarming rate. These seemingly incompatible facts and statements have prompted intense scientific debate and discussion in recent years.

Jack Dykinga/Nature Picture Library
Many attributes of nature that people value deeply have been severely deteriorated by our actions at the same time that biodiversity in many situations has not changed or has even increased. This fact is called the biodiversity conservation paradox. We can potentially resolve this paradox, at least in part, by considering that ecological patterns and processes can be different on small versus large spatial scales, an age-old topic of discussion in ecology. For example, the loss of roughly half the native bird species of New Zealand represents a global loss of bird biodiversity, given that all of these species were endemic, living nowhere else on the planet. The human-assisted introduction and establishment of roughly the same number of bird species as went extinct has maintained bird biodiversity in New Zealand but has done nothing to counter the global loss, given that all new arrivals already live elsewhere.
The fact that biodiversity changes differ across spatial scales, however, does not fully resolve the paradox. Most ecologists and conservation biologists—myself included—would still consider the New Zealand situation “bad” even if the regional losses were not also global losses. We place great value—entirely separate from scientific considerations—on wild nature, unsullied by human activities. In other words, human values influence how we describe and study biodiversity. Thus, to more fully resolve the biodiversity conservation paradox, we need to first examine how science and human values became intertwined in the development of the very concepts of biodiversity and of nonnative species. A collision of the dominant narratives on these two topics ultimately revealed logical inconsistencies—the paradox with which ecologists and conservation biologists have been grappling.
Biodiversity is good. This statement of value was declared as one of the postulates of the fledgling field of conservation biology in the 1980s, and by the 1990s my fellow students of ecology and I uncritically accepted it. But the impulse to preserve wild nature has much earlier origins. We felt a deep connection with nature and its stunning diversity of life forms, and we were eager to accept and promulgate arguments about the value of biodiversity for human well-being. Passionate pleas for biodiversity conservation from the likes of Norman Myers, Paul Ehrlich, and E.O. Wilson were codified in conservation biology textbooks, leaving no room for questions about whether biodiversity was good and whether human activities were bad for its maintenance. Habitat destruction and fragmentation, climate change, nutrient pollution, and nonnative species are bad, bad, bad, and bad. If anyone was talking about exceptions or nuances, they weren’t doing so loud enough for it to reach the classroom and the next generation of ecologists and conservation biologists.
With the rise of conservation biology, the concept of biodiversity came to sit at the core of conservation policy across the world. As of 2016, 196 countries or political units had ratified the international Convention on Biological Diversity (CBD), first opened for signing in 1993. (The United States is a solitary exception, having signed but not ratified the convention.) The first objective of the CBD is simply “the conservation of biological diversity,” and individual countries have pursued this objective in various ways, perhaps most often by providing legal protection to species at risk of extinction. The United States has its Endangered Species Act, Canada has its Species At Risk Act, and New Zealand has its Conservation Act.
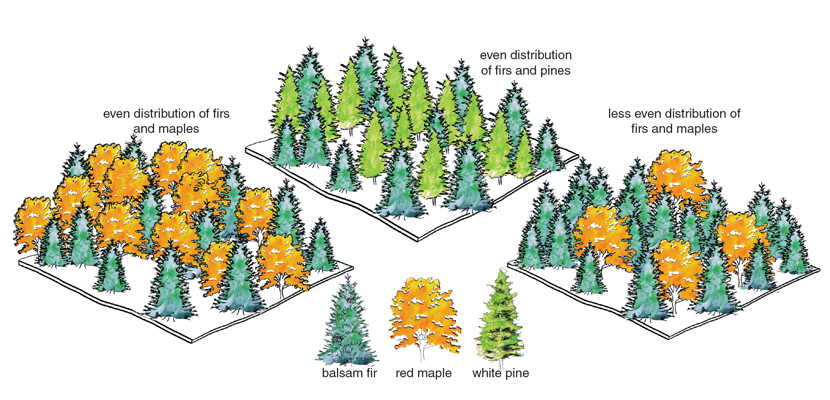
Illustration by Bethann G. Merkle.
Since the term biodiversity became widespread only in the late 1980s, ecologists have also been busy with the more mundane task of developing and calculating old and new indices to quantify it. The most straightforward index is species richness: the number of species in a given place and time. Indices of evenness aim to reflect the idea that, for example, a community with two equally abundant species is more diverse than one in which one of the species is dominant and the other very rare. Still other indices aim to capture phylogenetic or functional trait diversity, whereby a forest with both pine trees and maple trees is considered more diverse than one with only two species of pine (or maple) trees (see the figure at left). Species richness is the only metric that has been reported often enough, and in a sufficiently standardized way, to allow general comparisons across different regions, habitats, taxa, or scales of space and time.
Quantitative indices of biodiversity possess the scientifically useful attribute of incorporating no a priori value judgments about different species based on their geographic origins. If one native species goes extinct while one nonnative species becomes established (or vice versa), species richness is unchanged. These value-free calculations lie in stark contrast to the unambiguous incorporation of values into the study and politics of biodiversity, in particular with respect to nonnative species. In his pioneering text, The Ecology of Invasions by Animals and Plants, Charles Elton set the tone for invasion biology in 1958 that lives on to the present: “It is not just nuclear bombs and wars that threaten us…this book is about ecological explosions.” In other words, nonnative species, introduced from one continent to another, are bad; they must be prevented from establishing or be eradicated if established.
Graph Adapted from Sax and Gaines 2003. Spot illustrations by Bethann G. Merkle.
One reason given for deeming nonnative species bad is that they are an important cause of biodiversity loss. Flightless birds in New Zealand were not only prone to hunting by humans, but the smaller ones such as kiwis were (and are) also highly vulnerable to predation by introduced species such as possums and weasels. New Zealand’s Conservation Act distinguishes “indigenous” from “introduced” species, and one of its aims is to “control any introduced species causing damage to any indigenous species or habitat.” Indeed humans could be considered a nonnative species in New Zealand, responsible directly or indirectly for most, if not all, extinctions in recent centuries. But as we have seen already, nonnative species themselves contribute to local and regional biodiversity. If the case of New Zealand were unusual, we might consider it an exception to the rule that nonnative species cause biodiversity loss. It turns out, however, that the New Zealand story is quite typical.
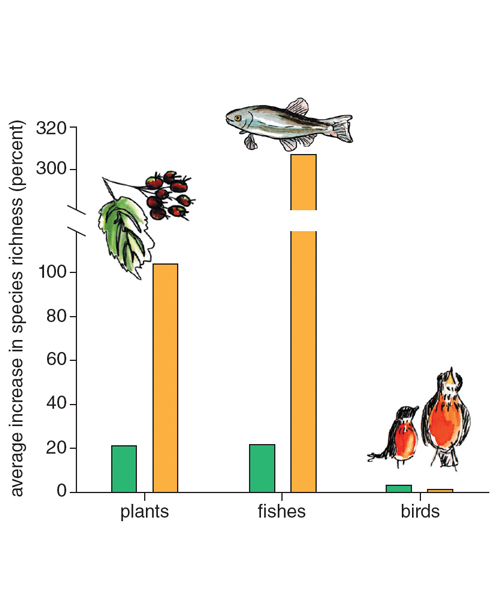
In the early 2000s, Dov Sax, now of Brown University, and his colleagues reported data that presented a major challenge to the conventional wisdom about nonnative species and biodiversity. In recent centuries, during which human impacts have been the most profound, oceanic islands ranging in size from less than 1 kilometer squared to greater than 250,000 kilometers squared have shown changes in species richness that mirror exactly the results in New Zealand. For birds, the establishment of one introduced species has matched every one of the many species that have gone extinct. For plants, very few species have gone extinct on these islands, and roughly the same number as originally present have been introduced and have established populations, thus doubling the original total. The species richness of freshwater fishes shows even more dramatic growth, with a fourfold increase over time on oceanic islands (see the figure at right). Ecologists are actively debating and studying whether these results imply a steady-state carrying capacity for some groups (for example, birds), a general “openness” to colonization in communities of some other groups (such as plants), or whether current diversity levels are transient, with future extinctions and colonizations altering or reversing the net changes seen so far.
In individual states or drainage basins in the United States (considered mainland areas, rather than islands), changes in species richness have been in the same direction, but of lower magnitude: no net change for birds, and a 20 percent increase for both plants and fishes, on average (see the figure just above). A 2009 study by Marten Winter, now of the German Centre for Integrative Biodiversity Research, and his colleagues showed a comparable increase of 20 to 25 percent for plants in European countries over the past 500 years. Since regional extinctions in these mainland areas very rarely represent global extinctions, we cannot reconcile this manifestation of the biodiversity conservation paradox by invoking the scale-dependence of biodiversity change. Species are being transported and introduced to new places around the world, so that regional diversity often increases, even as global diversity declines.
The net result of human activities in recent centuries thus appears on average to have been an increase, or at least no change, in species richness at the regional scale. If biodiversity is good, this news ought to be good, but one would be hard-pressed to find an ecologist who would say that. So, at the local or regional scale, perhaps it is not really biodiversity per se that we value.
Personally, I find value in the naturalness of a given ecosystem, flora, or fauna, with the benchmark for natural being the absence or at least minimization of human influence. I also have a love of variety in and of itself, which prompts me to seek out the unique, mysterious, and beautiful plants and animals in different corners of the globe. For example, the sight of scarlet and blue-and-yellow macaws flitting about on a riverside cliff in Peru, after waiting hours for fog to lift, is a profoundly thrilling experience. Of course, I consider the sustainable use of resources and promotion of public health to be major priorities for humanity, but regardless of whether biodiversity is the key ingredient in the maintenance of ecosystem services, I support its conservation because I want others to have opportunities to experience the sense of wonder and excitement in nature that inspires me. Ultimately, ecosystems—whether natural or intensely managed by people—provide the life support system for humanity. It is up for debate, however, how or whether biodiversity per se fits into this picture.
Graphs from Tilman et al. 1996; photograph courtesy of David Tilman.
For decades, environmental historians and conservation biologists have been discussing the reasons people value nature—and biodiversity. Some values are profoundly personal, such as feelings of moral obligation to protect nonhuman life or a spiritual connection with wild nature. For the most part, these values are outside the realm of science, except in the sense that we might quantify how many people share such values. There is nothing in the forest itself, however, that we scientists can measure to quantify its moral or spiritual value.
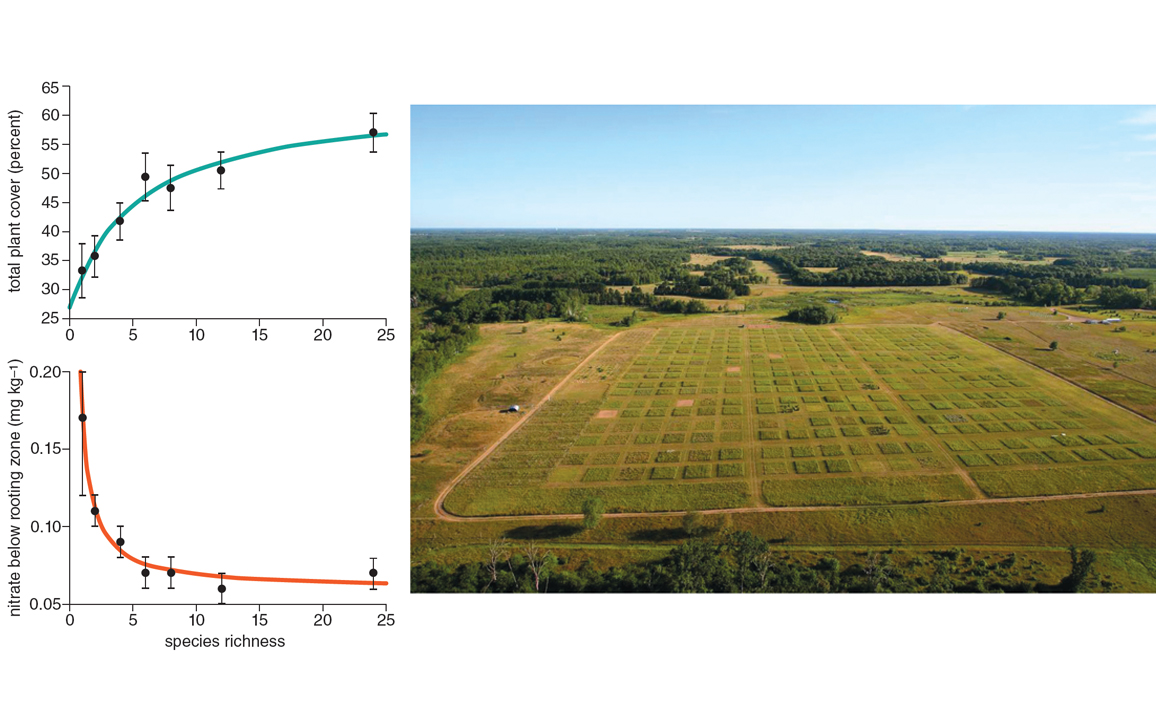
But there are some attributes of the forest we can measure that are of less ambiguous value to people, such as the filtration of the air we breathe or the water we drink. These attributes are referred to as instrumental values. A major thrust of ecological research over the past 25 years has been the experimental creation of ecosystems with different numbers of species and subsequent tests of how the number of species influences ecosystem functions such as biomass production by plants and the efficiency of nutrient use. These functions, in turn, influence the delivery of ecosystem services, such as the removal of carbon from the atmosphere and the reduction of fertilizer runoff and other waste nutrients leaching into water bodies. In the 1980s, Paul and Anne Ehrlich likened species extinctions to the loss of rivets from an airplane—a few popped rivets might have no effect, a few too many and the plane crashes. These studies effectively test this hypothesis.
Most often, biomass production and the efficiency of nutrient use do indeed increase as a function of the number of species in an ecosystem (see the figure above). Biodiversity enhances ecosystem function—not in every single experiment, but in most. This enhanced functioning would appear to provide proponents of biodiversity conservation with a powerful argument: We must conserve biodiversity because human well-being depends on it. Versions of this argument have made their way into documents from innumerable conservation organizations and government departments across the world. The website of the New Zealand Department of Conservation (http://bit.ly/2gg6vL4), for example, states that biodiversity “provides the services we rely on for our quality of life, our prosperity, and ultimately our survival.” The government of Québec, my home province, says essentially the same thing on its website (http://bit.ly/2gpCaNM): “Thanks to biodiversity, ecosystems can contribute to human well-being in terms of health, safety, and material comfort” (translated from French).
But wait a minute. We have already learned that nonnative species introductions have caused regional-scale biodiversity to increase rather than decrease in recent centuries. And if more species means better-functioning ecosystems, then the net effect of many nonnative species introductions and few native extinctions should be enhanced ecosystem function. Here the “biodiversity is good” argument collides with the “nonnative species are bad” argument, revealing an inconsistency.
Considering the link between biodiversity and ecosystem function prompts a return to the issue of how patterns of biodiversity can depend on spatial scale. The largest and best-known experiments on this topic have been conducted with plants in grasslands, using experimental plots of 100 meters squared at the large end. This scale is typically referred to as “local,” being orders of magnitude smaller than even the smallest of the islands included in the “regional” studies mentioned earlier. On one hand, conservation biologists often dismiss results from such small scales as irrelevant to policy or management issues, which often pertain to entire national parks, counties, or even countries. In this case, however, the causal link from biodiversity to ecosystem function involves small-scale interactions among plants, and so study plots are of an entirely appropriate scale.
On average, a plant of a given species grows to a larger size if its immediate neighbors are plants of different species rather than of the same species. The best-known example involves grasses and nitrogen-fixing legumes, such as clovers. Legumes fix atmospheric nitrogen via bacterial symbiosis and so grow well in nitrogen-poor conditions, adding nitrogen to the ecosystem. Grasses take up nitrogen efficiently and grow rapidly but potentially deplete the available resource. The two types of plants thus produce more biomass collectively than either can when growing alone. In dense grassland, plants are closely intertwined, separated one to the next by centimeters rather than meters, if they are separated at all. A grass plant benefits from a clover immediately adjacent, but it does not benefit measurably from a clover growing a kilometer or even 100 meters away. The plant–plant interactions within plots of a few meters squared have a big impact on both species.
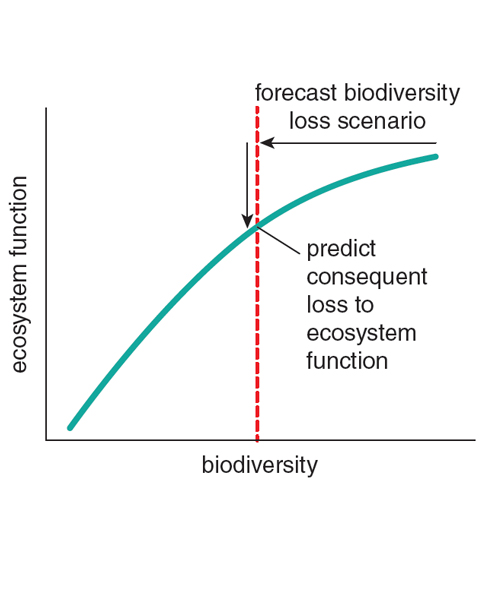
The fundamentally small-scale nature of ecological interactions that can create an effect of biodiversity on ecosystem function tell us that changes in biodiversity at a small scale are indeed of relevance to conservation issues. Plant biodiversity might be increasing regionally, but if grasslands or forests are experiencing net biodiversity losses locally, then the functioning of these ecosystems might be in decline. To accurately translate the results of experiments that test how biodiversity affects specific ecosystem functions into real-world implications, ecologists first must test the widespread assumption that local biodiversity is indeed declining (see the figure at right).
One scenario in which it is plain to see that local biodiversity has declined is when natural vegetation such as a tropical forest is converted to a crop monoculture or a parking lot. Agriculture in particular is the primary culprit implicated in the worldwide loss of natural habitat. But the link between biodiversity decline and ecosystem services in this case is not so simple. People convert natural vegetation to crop fields deliberately to maximize one ecosystem service: food production. Land conversion to agriculture can come at a steep cost to other ecosystem services, such as soil stabilization and water quality, and the use of crop mixtures rather than monocultures (which translates to greater biodiversity) can potentially contribute to strategies aimed at offsetting such costs. But how can we compare the relative value of a thousand bushels of corn versus less nitrogen in a stream? If you’ve got nothing to eat, the corn wins out; if you’ve got plenty to eat, maybe forest protection is the better strategy. In short, one cannot make a general statement about whether ecosystem services overall increase or decrease when a forest is converted to a crop field. Furthermore, any change in ecosystem function and services seems likely to have resulted more from plowing, leaving the field fallow each year, and the growth of annuals instead of century-old trees, than from a change in biodiversity.
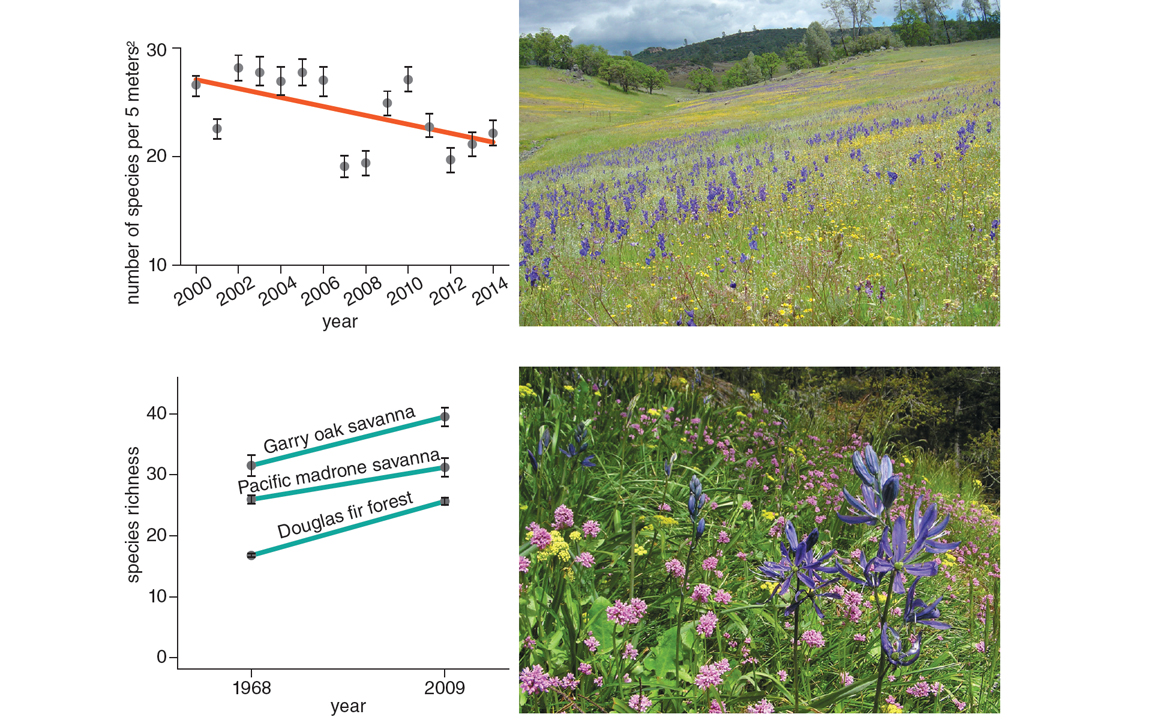
Graphs are adapted from Harrison et al. 2015 and McCune and Vellend 2013. Top photograph by Cathy Koehler, courtesy of Susan Harrison; bottom photograph courtesy of the author.
Outside of agricultural fields and parking lots, in ecosystems that have not experienced wholesale human conversion, the effects of introduced species, climate warming, and other environmental changes on biodiversity are much more variable. For example, in a California grassland, a decrease in rainfall—where soil moisture is already limiting to plants—appears to have caused a decline in local plant diversity over the past 15 years. In contrast, local plant diversity increased between 1968 and 2009 in forests and savannas of southern Vancouver Island as increased disturbance due to human development favored colonization by disturbance-adapted species, both native and nonnative (see the graph above). Because biodiversity-manipulation experiments do not involve habitat conversion, data from the kinds of studies just described are most relevant to assessing arguments about real-world causal links from biodiversity to ecosystem function.
Over the past five years, individual studies like these have been synthesized in several large meta-analyses. Across more than 200 studies that reported terrestrial plant species richness in local study plots for at least five years, a group of collaborators and I found that the average change in the number of species over time was statistically indistinguishable from zero (see the figure below). The same result was reported by Maria Dornelas of the University of St. Andrews and colleagues for 100 time series of species richness and other biodiversity metrics in a range of terrestrial and aquatic habitats from across the globe. A study by Robin Elahi of Stanford University and colleagues, focusing on the past 50 years specifically in marine coastal ecosystems, found an average net increase in local species richness in more than 400 time series, but an average net decrease in this measure among the subset of studies that experienced the greatest human impacts, such as sedimentation and pollution. All of these studies reveal immense variability from case to case, and at present we have only a limited understanding of the causes of so much variability.
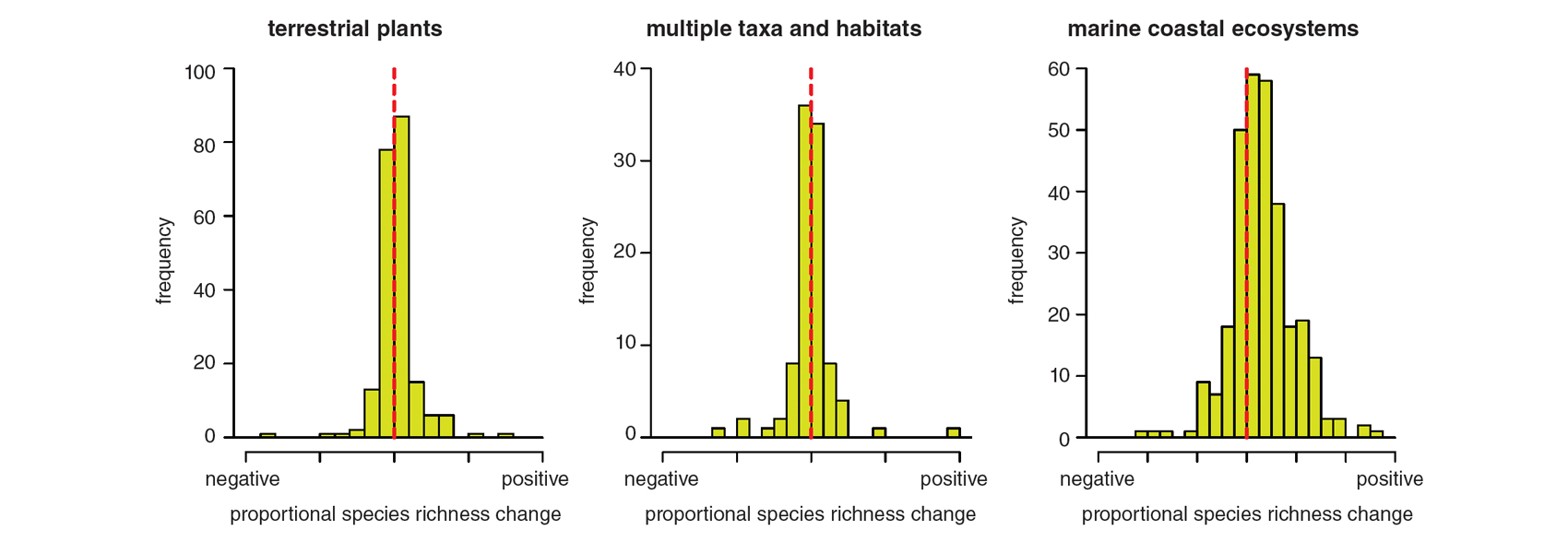
Data for the graphs are from Vellend et al. 2013, Dornelas et al. 2014, and Elahi et al. 2015.
The diversity-change meta-analyses, based collectively on many hundreds of studies and thousands of individual sampling sites, run counter to the assumption that local-scale biodiversity is generally in decline. The underlying data do come with caveats, such as uneven representation of geographic regions or levels of human impact. The same is true, however, of the experiments linking biodiversity to ecosystem function that form the other half of the scientific basis for the conservation argument (see the figure just above the “The Instrumental Value of Biodiversity” section). Overall, the results caution against overgeneralizing the argument that biodiversity decline in nature is a major cause of declining ecosystem services—an argument at the core of widely used justifications for biodiversity conservation. More generally, even if more data were to shift the means of these distributions slightly one way or the other, variability in the magnitude and direction of temporal trends, both at regional and local scales, deviates strongly from the general expectation of decline that forms the core of the biodiversity crisis narrative.
Although the absence of a “rule” of local biodiversity decline accompanying the global decline is seen by some researchers as strongly contrary to expectation, others find the results unsurprising. Elaborating on an expression from physics, the famous ecologist Joseph Grinnell stated in 1924, “Nature abhors a vacuum in the animate world as well as in the inanimate world.” He was communicating the idea that when ecological opportunities arise, for example when species are lost from an ecosystem, others are quick to exploit the opportunity. For example, when people create crop monocultures, it requires a tremendous effort to keep out unwanted weeds. Even in the experiments designed to test biodiversity-ecosystem function relationships, a major cost of research is the manual effort of keeping species out of plots where they aren’t supposed to be. If biodiversity is pushed down, Mother Nature often pushes back.
More generally, theory and some empirical data support the notion of a quasi-steady state in local biodiversity, with a tendency for deviations from the steady state to be followed by a return to the long-term average. Of course, it is possible for environmental change to alter the steady-state level, but predictions can go in either direction. For example, in a forest of eastern North America, one might expect climate warming to increase local plant diversity (forests to the south have higher diversity than those to the north), while anthropogenic nitrogen deposition strongly favors competitive dominance of one or a few species, thus causing local biodiversity to decline. Short of deliberately simplifying an ecosystem via the creation of a corn field or a strip mall, anthropogenically driven environmental change can push local and regional biodiversity in either direction.
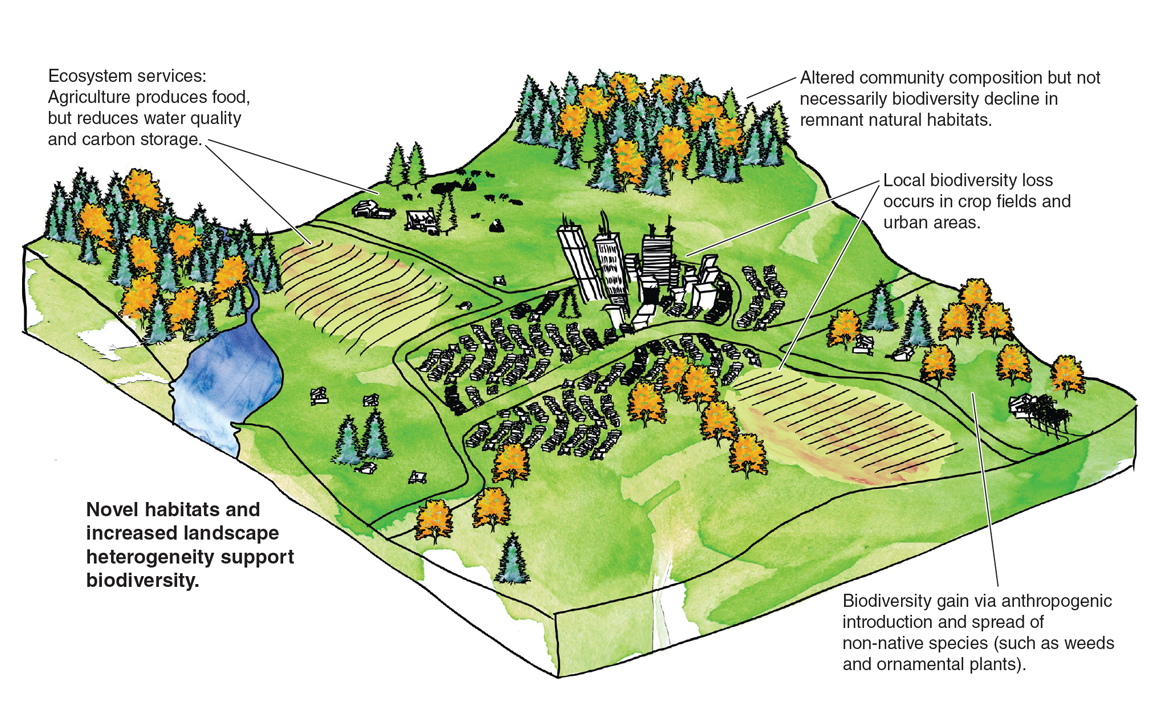
Illustration by Bethann G. Merkle
It is unsettling to have one’s view of the world called into question—in this case I had to face evidence that is contrary to the conventional wisdom in conservation biology imparted to me in the 1990s. Biodiversity is not generally declining at all spatial scales: Declines at the global scale are not generally seen at the regional scale and occur only in particular scenarios at the local scale. Human activities are not always to the detriment of biodiversity: Sometimes they are (for example, conversion of forest to crops), but sometimes they are not (for example, nonnative species doubling the numbers of plant species in New Zealand and on other islands). And nonnative species are not always the enemy, contributing sometimes in important ways to local and regional biodiversity as well as ecosystem services such as food and fiber production. The grasses and legumes that feed livestock in North America were mostly introduced from other continents (primarily Europe and Asia), as were the livestock themselves.
Returning to the biodiversity conservation paradox, I now think that perhaps its ultimate resolution lies in the fact that scientific definitions of biodiversity fail to capture what people really value about nature. If biodiversity is quantified in an objective way as some aspect of variety in an ecological community, then biodiversity can be high in a place that would be assigned low conservation value. On a visit to Hawaii, I recall seeing many kinds of birds in the lowlands, including northern cardinals, mourning doves, and house finches, all familiar species from my backyard in Canada, and all introduced to Hawaii from elsewhere, making them part of the local conservation problem. But if biodiversity is defined as whatever people value about nature, then science has nothing to contribute to the question of whether and how biodiversity has value. The definition provides a built-in answer.
To be clear, the argument here is not that all is well with the ecology of planet Earth. Even if biodiversity loss is found not in itself to be a major cause of compromised ecosystem services, there are plenty of serious ecological threats. The number of people on the planet and our per capita consumption of resources combine to make it highly uncertain that future generations will be able to enjoy the same level of well-being we enjoy today. This problem is clear after observing statistics on renewable and nonrenewable resources, and it is reflected in the eye-popping human impacts visible in images of the Earth taken from space. The temperature of the planet is rising, increasing the likelihood of detrimental heat waves, droughts, and coastal flooding. Many of the consequences of such changes for people flow through the biosphere in the form of altered crop production, fisheries yields, or natural water filtering.
What’s less clear is the degree to which biodiversity—by its scientifically justifiable definition—is a key player in mediating the consequences of global changes for human well-being. In the mountains to the north of Vancouver, British Columbia, where I used to live, forested watersheds are protected to provide clean water to the city’s residents. The intact forest is critical to the provisioning of clean water, but it seems unlikely that the number of tree species is so important. Indeed a typical forest stand in coastal British Columbia contains just a handful of tree species.
I suspect that scientists harbor deeper doubts about the validity of the utilitarian argument for biodiversity conservation than is evident in their published writings. In his 1996 book, The Idea of Biodiversity, David Takacs, then at Cornell University, interviewed many prominent conservation biologists and revealed a telling dichotomy between the reasons that inspire them to care about biodiversity, and the reasons they provide when trying to convince others to care. Inspiration often comes from an emotionally driven love of nature, and so, for example, the loss of endemic birds in New Zealand saddens us deeply. In contrast, when “selling” conservation, biologists more often employ the utilitarian argument that losing biodiversity will compromise human well-being, even if it’s not so clear that the transformation of New Zealand’s biota has been to the detriment of human well-being.
In short, a great burden was placed on the shoulders of the concept of biodiversity—to simultaneously represent all that we value about nature and to provide a way to quantify this value in a straightforward way. The concept might now be collapsing under this weight.
Saving the concept of biodiversity might require more clearly circumscribing the role of science in conservation. If, for example, an endangered species has no scientifically known or even plausibly hypothesized unique benefit to human well-being, we should be honest that the motivation for protection is driven by ethical values. It is important to note that such honesty requires acknowledging that the question of what society should do about a given situation (for example, a species at risk of extinction) is outside the realm of science, and so a scientist’s values carry no more weight than anyone else’s. In contrast, scientists do have a special standing when it comes to giving advice on how to achieve conservation policy or management goals and on predicting the possible consequences of action or inaction on a range of issues, including pollution, global warming, and land-use change.
In describing or predicting the consequences of anthropogenic environmental change, I think that scientific credibility can be safeguarded by keeping values out of the quantification of biodiversity as much as possible, or at least by clearly qualifying conclusions that incorporate values. I have been guilty myself of not clearly flagging value-driven decisions in the study of biodiversity. In my doctoral dissertation, I focused on how agricultural land-use history influenced forest specialist plants in New York State, and I described changes in “biodiversity,” despite deliberately having excluded species characteristic of open fields, such as those most likely to benefit from the environmental change. To a casual reader (meaning most readers), I thus ensured the appearance of an overall biodiversity decline, when really overall plant biodiversity may have shown an entirely different response.
I believe the example of my PhD research is representative of a great many similar studies, in which value judgments entered into the very calculations whose results might then appear to justify the values. Today, I would describe the decline I observed as a loss of forest specialist plants, not a decline in plant biodiversity, unless I had data that characterized the full set of plant species present over time and demonstrated a decline. That way, there would be far greater transparency regarding the values involved in developing any conservation practices based on this research. I worry that the conventional wisdom that was overturned by the studies of Dov Sax and others (but that still lives on to a considerable degree) was built on a foundation that included an awkward mixture of human values and data. In my experience, the presentation of any result that runs counter to the biodiversity crisis narrative is met with a chorus of responses from ecologists that start with “yeah, but…”. As the professional skeptics we are supposed to be as scientists, I think that ecology and conservation biology can benefit greatly if we consistently apply “yeah, but…” thinking not only to results that run counter to our personal values, but also to those that align with them. I have certainly started to do so myself.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.