Not Just Going with the Flow
By Frank Eliot Fish, George Lauder
Modern visualization techniques show that aquatic animals can modify their fluid environment to increase the efficiency of swimming and food collection.
Modern visualization techniques show that aquatic animals can modify their fluid environment to increase the efficiency of swimming and food collection.

DOI: 10.1511/2013.101.114
Swimming fish and dolphins appear to move effortlessly through the water. Even when they glide, they don’t seem to lose any speed. However, rigid and fixed laws of physics and principles of hydrodynamics dictate how water flows around an animal. This flow determines the forces the animals must generate and the energy they must expend to move. Animals propelling themselves through water must contend not only with pushing back on the fluid but also with forcing their way through an incompressible medium. Despite the inflexibilities of these physical forces, underwater animals have developed highly effective means to control flow and move economically through the aquatic environment.

John Dabiri.
The observation of the interaction between an aquatic animal and the water around it, called its flow field, goes back to Leonardo da Vinci. Da Vinci recognized the advantages of the streamlined shape of a fish based on the flow surrounding it. He argued that the fish could move through the water with little resistance, or drag. The streamlined shape allowed the water to flow smoothly over the body. Later, in 1809, Sir George Cayley, the father of aerodynamics, examined the streamlined body shapes of a trout and a dolphin. He considered their similar shapes to be solids of a design that offered the least resistance to flow. More recently, Steven Vogel of Duke University introduced biologists to the interaction of organisms and flow with his 1981 book Life in Moving Fluids. His contribution resulted in a greater appreciation of the interaction between animals and the liquid medium.
Manipulation of flow is accomplished both passively and actively. Animals use passive mechanisms involving design of the body and texture of their surfaces, which alter flow conditions against the body surface in order to reduce drag. On the other hand, active control of flow involves mobile fins and paddles, which regulate water movements that are shed into the wake as vortices. Vorticity is the tendency of a fluid to rotate or spin. An extreme version of vorticity is a vortex. The vortex is a spinning, cyclonic mass of fluid, which can be observed in the rotation of water going down a drain, as well as in smoke rings, tornadoes and hurricanes.
Up until the last 20 years, the control of flow by animals has largely been conceptualized rather than visualized. Expectations of the interaction of animals and water flow were based on simple engineered systems. Streamlined forms, synthetic wings, wavy plates and polished surfaces were the standards to compare with animals. However, animals exhibit a wide diversity and complexity of shapes and movements, which can influence the flow dynamics in ways not previously envisioned. Animals change shape during motion, unlike human-designed systems, further complicating analyses of motion in a fluid environment.
Biologists have recently been employing the same techniques as engineers to visualize flow, and these methods have greatly aided biologists in describing and quantifying the flow fields around animals. One mechanism is to introduce dye into the water along the surface or in the wake of a swimming animal, which shows a continuous record of the trajectory of the fluid. Similarly, particle image velocimetry (PIV) can visualize the pathway of the flow by illuminating reflective particles introduced into the fluid. The particles are illuminated with a laser that has been projected into a wide “sheet” rather than a single beam. This optical method also can define the velocity of the flow in a two- or three-dimensional field around the animal. Tracking the particles in the flow requires high-speed cameras and sophisticated computer software. Compared to these experimental methods of flow visualization, computational fluid dynamics uses computers to simulate the flow from numerical solutions, which are based on theoretical equations governing fluid movement.
Research using these varied flow visualization techniques has given new insights into how animals manipulate flow. Long-standing ideas are being tested regarding the best designs and mechanics of movement with regard to enhanced propulsion, reduction in drag, and the coordination of feeding and locomotion. Understanding how animals can control flow has immense implications not only for understanding the evolution of aquatic species, but also for developing biologically inspired machines and even for elucidating global climate change.
Whether jellyfish or fish, minnows or whales, swimming animals propel themselves by producing a thrust force in opposition to a resistive drag force. Thrust and drag are the yin and yang of hydrodynamics. Under conditions of a constant swimming speed, thrust and drag must balance each other. To swim fast, animals need to be able to minimize drag and maximize thrust. The amount of thrust and drag generated has ramifications on the total energy cost of swimming. As the animal pushes against the fluid medium to propel itself, it transfers kinetic energy from its body motions to the water. In addition, drag consumes energy from the movement of the body, which decelerates the animal unless it is countered by propulsive motions produced by muscular actions.
Drag is produced mainly from frictional and pressure forces resulting from the interaction of the fluid and the body. Frictional drag is caused by friction between the skin and a thin layer of fluid in close proximity, called the boundary layer. The fluid touching the skin adheres to it without slipping due to the viscosity or “stickiness” of the fluid. Because of this no-slip condition, the friction due to viscosity shears the flow within the boundary layer. This shear is like pushing on the top of a deck of cards lying on a table: The bottom card stays fixed in position while the top card moves with the hand and all the cards in between are displaced slightly. The boundary layer possesses energy that maintains its flow against the skin. However, the frictional shear within the boundary layer represents energy lost as drag.
Fish can secrete mucus or slime over the body to reduce frictional drag. The slime is a combination of lipids and proteins, many of which contain long chains of molecules, and some of which can act as surfactants (lowering the surface tension of a fluid). Since the late 1940s, engineers worldwide (working on ships, submarines and pipes) have established that the addition of dilute solutions of long-chain polymers into flow is an effective means of drag reduction. The slime similarly reduces the viscosity of the water around the fish. For instance, the barracuda, Sphyraena argentea, possesses slime that reduces frictional drag by as much as 66 percent, and the fish can reach speeds of up to 27 miles per hour in short bursts.
The other type of drag, pressure drag, is dependent on the shape of the body in a flow. The hydrodynamic association between pressure and velocity of a fluid is described by the Bernoulli principle. First published in 1738 by the Swiss mathematician Daniel Bernoulli, the principle states that pressure and velocity are inversely related, so that as velocity decreases, pressure increases and vice versa. The impact of the water on the face of the body decelerates the flow, which increases the pressure. (Think about how your hand is pushed backward when you hang it out of the window of a moving vehicle.) As the water follows the contours of the body, it accelerates and thus reduces the pressure. Before being ejected into the wake, the flow decelerates again with an increase in pressure, but not to the same extent as at the front of the body. The difference in pressures around the body generates an imbalance in forces and results in the pressure drag. The physical manifestation of the pressure drag is the width of the wake: Narrow wakes represent less pressure drag than broad wakes. Broad wakes are due to the interaction of the pressure changes and the boundary layer. If the energy within the boundary layer flow is insufficient to maintain a downstream direction of flow, the pressure changes will cause the boundary layer to prematurely separate from the body surface, forming eddies, increasing vorticity and producing a broad wake. Thus the drag is minimized particularly when fluid moving along the body surface remains attached.
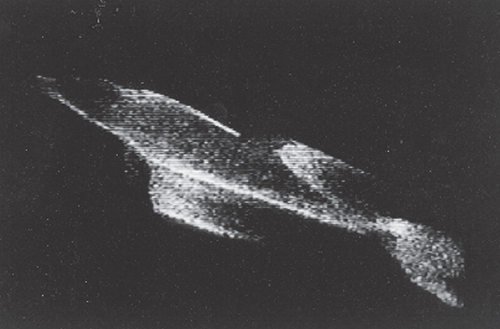
James Rohr
Flow is controlled mainly by streamlining the body shape to minimize drag. It is no accident that fish, dolphins and even submarines have an elongated, teardrop design. This shape gives the lowest drag per volume. Pressure differences are minimized and the boundary layer remains attached, keeping the wake narrow. Jim Rohr and Michael Latz at the Space and Naval Warfare Systems Center demonstrated this fact for gliding dolphins. These researchers were able to visualize the flow around the dolphins using bioluminescence from marine phytoplankton. Shear stresses within the flow around the body perturbed the unicellular plankton and caused it to light up.
Although it might intuitively seem that a smooth surface will help minimize drag in swimming animals, a rough skin can actually be more effective in controlling flow over the body and reducing drag. A textured surface can generate turbulence within the boundary layer region near the body surface. Although this turbulence increases the frictional drag, it infuses more energy into the boundary layer. The energy added by turbulence stabilizes the boundary layer by allowing it to overcome adverse pressure changes. The turbulent boundary layer is thus less likely to separate and increase the pressure drag. The same formation explains why golf balls with dimples travel faster and farther than smooth balls. The dimples cause turbulence near the ball surface and this increased energy delays flow separation and narrows the width of the wake.
Fish have a variety of complex structures on the body surface that may act like golf ball dimples. Swordfish, Xiphias gladius, have an elongated rostrum (or snout) with a rough surface of craters and bumps, which may act to induce turbulence in the boundary layer over the body. Many bony fish are covered with small ctenoid scales (named for the small tooth-like projections on the scales’ posterior, downstream edge) that could also alter boundary layer flow, although as yet very little research has been done on the hydrodynamic effect of bony fish scales.

Denticle photos courtesy of Johannes Oeffner; shark photo courtesy of George Lauder.
On the other hand, the skin of sharks has been studied for many years, and has a remarkable surface structure. Shark skin is covered with numerous small denticles (or scales) that typically have three ridges on the surface and downstream-facing prongs. These closely packed denticles extend above the skin surface and into the boundary layer region. Each denticle is like a small tooth—with enamel, dentine and a pulp cavity in homology with mammalian teeth. Different shark species have varying details of denticle structure, and separate regions of the body display denticles with specialized shapes. Denticles near the head can have a flat paver-like surface and reduced ridges, whereas those on the body and tail possess long prongs and deep ridges.
Although there has been considerable research by engineers on the drag-reducing function of rough materials attached to a rigid surface, it is important to study the function of denticles when the skin surface is moving as it does during locomotion by live sharks. Because experiments to study shark skin hydrodynamics are not easily conducted on live fish, a robotic device instead can be used to move isolated pieces of shark skin in a laboratory setting. One of us (Lauder) has designed such devices to allow flexible sections of skin to “swim” in a recirculating flow tank, which mimics the way that the skin moves on a live shark. Patterns of water flow over the skin pieces can be measured and compared to skin that has had the denticles removed.
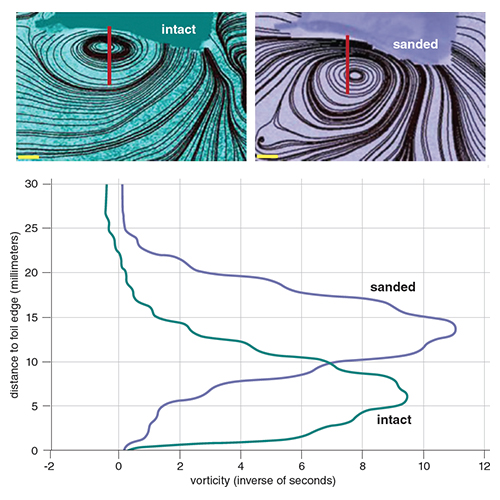
Images courtesy of Johannes Oeffner and George Lauder. Graph by George Lauder and Tom Dunne
Such studies have revealed that shark skin denticles enhance swimming speed, as skin pieces swim more slowly after the denticles have been removed to create a smoother surface. The increase in swimming speed when denticles are present may be the result of drag reduction, as the denticles function to generate a turbulent boundary layer. But experimental data on water flow over shark skin also suggest that the denticles alter flow in a way that might enhance thrust. Imaging flow over flexible membranes made of shark skin shows that a vortex is formed adjacent to the surface. This vortex contains a low-pressure region that acts to enhance the thrust produced by the skin. When denticles are removed, the leading edge vortex moves farther away from the surface, and the effect of the low-pressure suction on propulsion is lessened.
Propulsion requires that animals generate thrust by accelerating a mass of fluid, which can be accomplished by the actions of the body itself, from appendages such as fins and paddles, or by ejecting the fluid in jets. The accelerated fluid is configured as vortex rings with a central jet—the same formation as blowing smoke rings. Indeed, John Dabiri of the California Institute of Technology and Ian Bartol of Old Dominion University in Virginia used dye injection and PIV to visualize the pulsatile ejection of fluid in the wake of jellyfish and cephalopods (such as squid and octopus), respectively. The propulsive actions of jellyfish and squid produce a series of distinct vortex rings with the jet oriented directly behind the animal, similar to smoke rings.
Adapted from Lauder and Drucker, 2002, by Tom Dunne.
The wriggling body of a fish produces much of the vorticity that is shed into its wake. The action of the tail as it reaches its maximum sideways deflection rolls the vorticity up into a vortex. As the vortex drifts away for the fish, a new vortex forms with an opposite spin direction as the body flexes and the tail reverses direction. The alternating vortices are linked three-dimensionally. The continuous lateral undulations of the body and tail organize the wake as a staggered array of interconnected vortex rings. These vortex rings induce a jet flow that is oriented to the side and backward and laced through the center of the rings. The jet flow produces thrust to overcome the drag of the body.
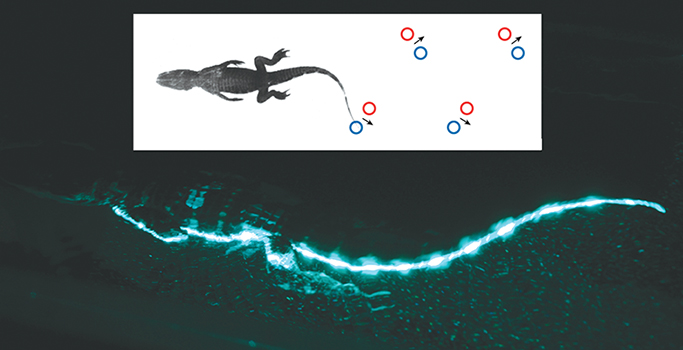
Courtesy of the authors
When we examined swimming of juvenile alligators using PIV, a different pattern of vortices in the wake emerged. The alligators swim by undulations, as waves were passed down the body and long tapering tail with increasing amplitude toward the tail tip. Vortex rings were shed from the tail at the end of each half-stroke. Each vortex ring was elongated as fluid was entrained into the wake by the whip-like action of the tapering tail. In addition, each vortex ring was not linked to the vortex ring generated from the proceeding half-stroke, as was observed for fish.
Optimal thrust propulsion and efficiency can be achieved by controlling the pattern and periodicity of the vortices in the wake. Production of an effective jet flow in the wake of a fish is possible within a narrow range of what is called a Strouhal number, a measure that represents the degree of unsteadiness in the flow, which dominates the wake dynamics. This nondimensional parameter is the product of the propulsive frequency and lateral displacement of the caudal (or tail) fin, which produces the vortices, divided by the velocity of the animal. The Strouhal number has maximal efficiency in the range between 0.2 and 0.4. Fish and cetaceans often remain within this optimal range and in so doing achieve propulsive efficiencies higher than standard marine propellers. Alligators, however, have Strouhal numbers above the optimal range. Efficiency and propulsive performance may also be diminished in this amphibious species compared with more specialized swimmers.
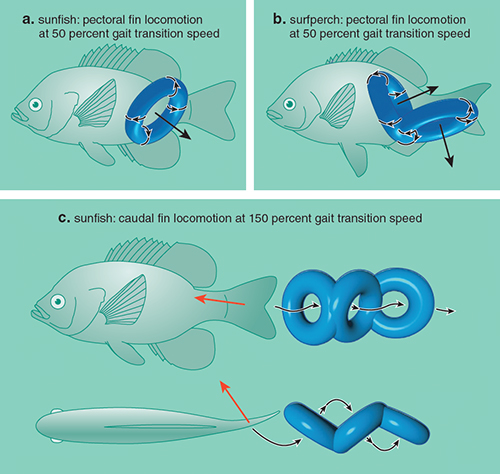
One of us (Lauder) used wake visualization techniques to reveal that the pattern of vortices is associated with gait changes by swimming animals. These gait changes are similar to the transition of gaits observed in terrestrial locomotion. Gait changes in aquatic animals occur with different speeds, bouts of acceleration and maneuvering.
Work performed on bluegill sunfish shows that these fish use different combinations of fins for propulsion, depending on swimming speed. At slow speeds, bluegill sunfish use only their paired pectoral fins (located on each side of the fish near the head) to generate thrust, whereas at higher speeds these fish recruit additional fins (caudal, anal on the underside and dorsal on top) to generate larger propulsive forces. The change in gait reflects constraints on thrust production by the pectoral fins. These fins generate a double-linked vortex ring with a central jet oriented to the side. This swimming gait promotes control and equilibrium at low speeds. As speed increases the jet is oriented more laterally. This increases stability, but limits thrust production. The change to unpaired fins and body undulations permits more momentum to be transferred from the fish to the water in the downstream direction. In contrast to gait changes in bluegill sunfish, surfperch use a different strategy as speed increases, and are able to continue to use their pectoral fins through a wide range of swimming speeds without recruiting other fins. As surfperch swim faster, the flapping motion of their pectoral fins allows reorientation of the pectoral vortex rings to direct the central jet flow directly downstream and hence generate thrust.
The generation of vorticity is an inevitable consequence of propulsion in a fluid medium. The vorticity shed into the wake represents a substantial loss of energy for swimming animals. But animals can swim more efficiently if they are able to extract energy from the swirling vortices. As energy is a limited resource that can affect the survival and reproductive efforts of an animal, there are adaptive benefits to having hydrodynamic mechanisms that recapture energy destined to the entropy of the wake.
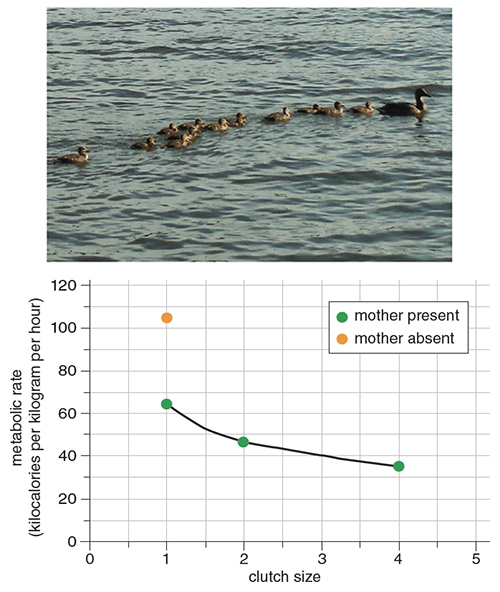
Data courtesy of Frank Fish, graph by Tom Dunne. Photo from Wikimedia.
Aquatic animals often travel in highly organized formations such as schools. By aligning themselves in a defined pattern, individuals in the group can take advantage of flow patterns generated by others to reduce drag and enhance locomotion performance. Vortices generated by leading individuals pass backward and impact trailing individuals. If a trailing animal is oriented parallel and moving in the same direction to the tangential velocity of the vortex, the body will experience a reduction in its relative velocity, which reduces drag. Vorticity is shed into the wake of a passive body as two rows of counter-rotating vortices (which is called a Kármán vortex street, named after Theodore von Kármán, a pioneer in aerodynamics), where the optimal position for drag reduction is directly behind the leading body. One of us (Fish) was able to train ducklings to swim behind a duck decoy in a flow tank. The ducklings would swim in single file. Metabolic measurements showed that as the number of ducklings in the line increased, the energetic cost per duckling decreased. This effect was more pronounced with the presence of the decoy, generating a wake. Furthermore, it was determined that last duckling in the line swam with the least effort. By swimming behind their mother to reduce locomotion effort, ducklings act just like stock car racers and bicycle pace lines.
One of our (Lauder’s) doctoral students, James Liao, swam fish in a flow tank behind a cylindrical object. Fish could maintain position behind the cylinder, which produced a vortex street. By slaloming between the vortices while staying inside the wake of the object, a fish can reduce its drag and even get a push forward. Trout can swim in between vortices with very little activity of body musculature. This reduction in muscle activity when swimming strongly suggests that fish are experiencing a considerable energy savings compared to free-stream locomotion.
Although similar in pattern to the Kármán vortex street, the vortices produced by the wagging tail of a swimming fish spin off with an opposite rotation. This is a thrust-type vortex system. In this system, the momentum jet is pointed directly behind the fish. An animal immediately following the fish would experience an increased flow velocity with its concomitant increase in drag. Daniel Weihs of Technion-Israeli Institute of Technology determined that the optimal configuration of a school of fish is a diamond pattern with the trailing fish outside the vortex street. In this position, the trailing fish takes advantage of the spin of the vortex. As the leading fish leaves the two rows of vortices in its wake, the spinning fluid has a forward-directed component on the side away from the fish. Trailing fish use the forward-directed velocity of the vortex by swimming laterally to the vortex street. As the trailing fish is swimming in the same direction as the tangential velocity, its relative velocity is less than the swimming speed of the school, so the individual fish experiences a reduced drag. Fish in a diamond configuration experience a reduction of the force generated for swimming by a factor of four to six.
Small cetaceans often position themselves beside and slightly behind the maximum diameter of a larger animal. In this position, the smaller individual gains an energetic benefit. This effect is vital particularly for young dolphins to maintain speed with their mothers. The flow that is channeled between their bodies induces an attractive force due to the Bernoulli effect, which draws the infant along with the mother. Weihs estimated that a neonatal dolphin could use this mechanism to gain up to 90 percent of the thrust needed to move alongside its mother. Although the young gain a benefit, the larger mother will experience increased drag from her towed offspring.
Every free-swimming animal requires flow manipulation to satisfy two opposing functions: stability and maneuverability. Stability acts to self-correct for disturbances and maintains a desired postural attitude. Maneuverability does the opposite by allowing a controlled instability to create a change in direction, as well as enabling the animal to stop and start. Swimming animals, which are suspended in the water without a solid support to lean against, must be plastic enough to do both, but with an economy of action so as not to uncontrollably destabilize their entire system. Morphological additions can control the pattern of flow for improved stability and maneuverability.

Ian Bartol
Bartol and his colleagues examined the boxfish, which is encased in a rigid, bony carapace. This form compromises stability control, as the fish is incapable of flexing its body and must instead use combinations of movements by its fins. These extra movements would drain the fish of energy just to stabilize the body and hold position in a water current. However, the carapace is equipped with structures called keels that aid in passively stabilizing the body. The keels are located dorsally and ventrolaterally (on each side of the belly). Vortices are generated as water flows past the keels. These vortices generate suction forces that increase as the fish is angled to the flow. The forces return the body to a more stable orientation.
Although stability is important to swim straight ahead or hold position in a flow, maneuverability may be more important as animals rarely move in straight lines. When life and death are on the line, complex movements are needed for prey to outmaneuver a predator, or for a predator to turn fast enough to catch elusive prey. One of us (Fish) has focused on the hydrodynamics associated with the aquabatic maneuvers by the humpback whale. These maneuvers are required to catch fish that are many times smaller and more maneuverable than the whale. Humpback whales use their elongated pectoral flippers, which can be up to a third of the length of the body, to execute tight turns and corral fish.
Just like an airplane performing a banking turn, the flippers generate a lift force that turns the whale. The magnitude of the lift is correlated with the angle at which the flippers meet the oncoming water flow (called the angle of attack). The lift will increase until the angle of attack reaches a critical point known as stall, where there is a dramatic loss of lift. On an airplane, stall could be catastrophic, but for a whale buoyed up by the water, stall would mean the inability to complete a turn and loss of food. In effect, stalling the flipper during a turn would be like driving along a curved road and hitting a patch of ice; the car would shoot off the road moving tangentially to the turn.

Top image courtesy of William Rossiter; bottom two images courtesy of Eric Paterson.
Delaying stall at high angles of attack becomes critical for the humpback to make tighter turns and catch a meal. For this purpose, the whale flipper has a novel design. The leading edge of the flipper has 10 prominent, rounded bumps or tubercles. Laurens Howle, an engineer at Duke University, and his colleagues tested idealized models of flippers with and without tubercles in a wind tunnel at the U.S. Naval Academy. They found that the flipper with tubercles stalled at higher angles of attack than the flipper with a straight leading edge. Howle and his colleagues also demonstrated that the tubercles enhanced lift production, and the hydrodynamic benefits occurred without an increase in drag.
To understand why the tubercles work, it helps to first understand normal flow around a winglike structure. Flow impacts the leading edge of a wing first, and then it is deflected straight rearward, both above and below the wing surface. As the wing is canted up with a positive angle of attack, the flow produces a low pressure over the upper surface because the fluid moves faster than the flow over the lower surface, which is deflected downward. The fluid on the upper surface flows faster because it must travel farther in the same amount of time that it takes the lower flow to travel from leading to trailing edges. Both upper and lower flows must meet at the trailing edge at the same time to maintain continuity of the flow. This idea was first postulated by da Vinci. Lift is produced as a consequence of the pressure difference between the two surfaces and the momentum of the deflected fluid. As the stall angle is approached, the flow over the upper surface does not have enough energy to hug the surface and starts to detach. The eddies that form over the upper surface at stall disrupt the pressure difference and reduce the lift.
Eric Paterson, an aerospace engineer now at Virginia Polytechnic Institute and State University, and his student, John Reifenberg used a computational fluid dynamics model to illustrate how flow is modified by the tubercles. The computational model showed that as the flow moved directly over the tubercle it was accelerated toward the trailing edge of the wing. In effect, the flow was energized, which kept it attached to the wing surface and prevented stall.
How was the flow over the tubercles energized? That result was explained by the flow in the troughs between the tubercles. As the flow impacts the leading edge in the troughs between two adjacent tubercles, it is deflected into the center of the trough, producing a pair of vortices with opposite spins. Each vortex that is immediately flanking the flow over the tubercle has a spin, which is in the same direction as the flow. Sandwiched between two vortices, the flow over the tubercle is energized and accelerated, which avoids separation from the wing surface and prevents stall. This action is analogous to a baseball pitching machine, which accelerates a ball by squeezing it between two counter-rotating wheels.
The advent of modern techniques of flow visualization and computational hydrodynamics has given biologists greater insight into the dynamics of flow around swimming animals. Physical adaptations associated with movement in an aqueous medium can be investigated from functional, ecological and evolutionary perspectives. Even problems with global implications have benefited from a greater understanding of bio-fluid dynamics. Dabiri and Kakani Katija Young of Woods Hole Oceanographic Institution have proposed that the flow patterns produced by the locomotion actions of organisms can be credited with ocean mixing. In other words, the constant motion of animals can serve as an efficient mechanism of vertical energy transport in the oceans. The cacophony of flows induced by the multitude of swimmers produces a biogenic turbulence that stirs the ocean. The vorticity shed by animals into their wakes increases the energy for mixing, which can affect global ocean circulation and facilitate the transport of nutrients. The implication of this energy transfer is that swimming animals may be a component modulating global climate change.
Our research programs have focused on the ability of animals to manipulate flow, which have presented new opportunities in the development and improvements of advanced technologies. In collaboration with engineers and physicists, we are involved with the emerging fields of biomimetics and bioinspired design. These fields attempt to incorporate novel structures and mechanisms from nature into the design and function of engineered systems. Insight into different mechanisms related to the control of flow can be used to create and improve everyday technologies. Based on the flow dynamics of fish schools, the placement of turbines in a modern wind farm can similarly extract greater amounts of energy. Lift generation by control surfaces for marine vessels could have enhanced maneuvering performance by the addition of leading edge tubercles based on the humpback whale flipper.

Top right image courtesy of Hossein Haj-Hariri; photographs courtesy of George Lauder.
Currently, we are investigating the feasibility of flow control by the flexible, oscillating fins of the sunfish, as well as manta rays and stingrays, for biomimetic application to robotic systems, specifically toward improvement of autonomous underwater vehicles (AUVs). AUVs are used to survey and sense the ocean environment. The inclusion of locomotion specializations from animals into AUVs could manipulate flow to reduce energy expenditure, increase thrust, provide high efficiency and enhance maneuverability. Both organism and machine encounter the same physical forces, as animals and engineered systems must function in a fluid environment that is 800 times denser and 60 times more viscous than air, and where drag is a major hindrance to movement. We expect that the development of the next generation of AUVs will include propulsion by flexible, oscillating fins. Engineering has given us sophisticated imaging methods that allow us to uncover the secrets that aquatic animals use to gain advantage in their fluid environment, and that biological information can now be fed back into the advancement of engineered systems.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.