How the Retina Works
By Helga Kolb
Much of the construction of an image takes place in the retina itself through the use of specialized neural circuits
Much of the construction of an image takes place in the retina itself through the use of specialized neural circuits

DOI: 10.1511/2003.11.28
The retina is a filmy piece of tissue, barely half a millimeter thick, that lines the inside of the eyeball. The tissue develops from a pouch of the embryonic forebrain, and the retina is therefore considered part of the brain.
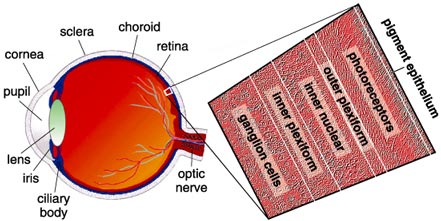
This most important part of the eye has a basic structure similar to that of a three-layer cake, with the bodies of nerve cells arrayed in three rows separated by two layers packed with synaptic connections. The retina includes both the sensory neurons that respond to light and intricate neural circuits that perform the first stages of image processing; ultimately, an electrical message travels down the optic nerve into the brain for further processing and visual perception.

Helga Kolb
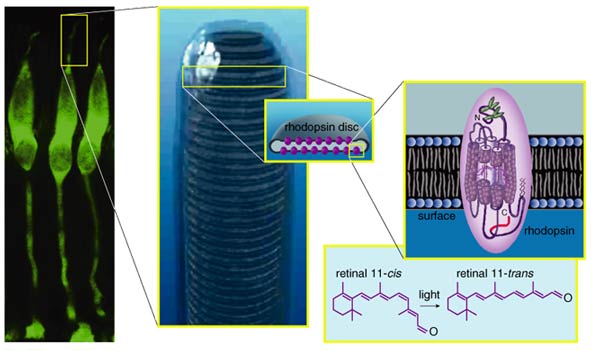
Intuitively, one might expect that the surface of the retina (the layer exposed to the liquid in the eyeball's vitreous chamber) would contain the sensory cells, the photoreceptors, but actually these cells lie at the very back of the retina; light rays must pass through the entire retina before reaching pigment molecules to excite. This is because the pigment-bearing membranes of the photoreceptors have to be in contact with the eye's pigment epithelial layer, which provides a steady stream of the vital molecule, retinal or vitamin A. Retinal becomes fixed in the photoreceptors' opsin proteins, where this small molecule changes its conformation in response to photons, or packets of light. Once retinal molecules are exposed to light and undergo their conformational change, they are recycled back into the pigment epithelium. This tissue behind the retina is usually very dark because its cells are full of melanin granules. The pigment granules absorb stray photons, preventing their reflection back into the photoreceptors, which would cause images to blur. They also protect the cells from too much exposure to light radiation.
All vertebrate retinas contain at least two types of photoreceptors—the familiar rods and cones. Rods are generally used for low-light vision and cones for daylight, bright-colored vision. The variations among animal eyes reveal adaptations to the different environments in which they live. Most fish, frog, turtle and bird retinas have three to five types of cones and consequently very good color vision. Keep in mind that reptiles and fish are "cold blooded" and need to be active in the warm daytime. Most mammals have retinas in which rods predominate.
Helga Kolb
When the number of mammals started to explode as the dinosaurs died out, the Earth was likely a dark place covered in ash and clouds; the tiny, fur-covered early mammals were able to generate their own body heat and developed visual systems sensitive to dim light. Modern rodents such as rats and mice, which are nocturnal animals, still have retinas overwhelmingly dominated by rods; their cones are small in size and only make up 3 to 5 percent of their photoreceptors.
Tom Dunne
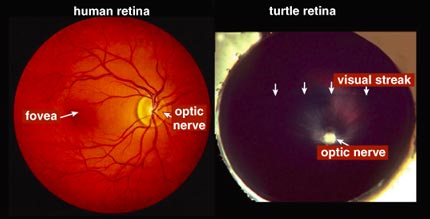
Most other mammalian retinas also have a preponderance of rods, and the cones are often concentrated in specialized regions. In species such as cats and dogs, images focus to a central specialized area, aptly called the area centralis, where cones predominate. The retinas of mammals such as rabbits and squirrels, as well as those of nonmammals like turtles, have a long, horizontal strip of specialized cells called a visual streak, which can detect the fast movement of predators. Primates as well as some birds have front-projecting eyes allowing binocular vision and thus depth perception; their eyes are specialized for good daylight vision and are able to discriminate color and fine details. Primates and raptors, like eagles and hawks, have a fovea, a tremendously cone-rich spot devoid of rods where images focus.
Nicolas Cuenca and Helga Kolb; Carlos Rozas; Cuenca and Kolb
Primates, in fact, have what is called a duplex retina, allowing good visual discrimination in all lighting conditions. The fovea contains most of the cones, packed together as tightly as physically possible, and allows good daylight vision. More peripheral parts of the retina can detect the slightest glimmer of photons at night. Most mammals have two types of cones, green-sensitive and blue-sensitive, but primates have three types—red-sensitive as well as the other two. With our cone vision, we can see from gray dawn to the dazzling conditions of high noon with the sun burning down on white sand. Initially the cone photoreceptors themselves can adapt to the surrounding brightness, and circuitry through the retina can further modulate the eye's response. Similarly, the rod photoreceptors and the neural circuitry to which they connect can adapt to lower and lower intensity of light.
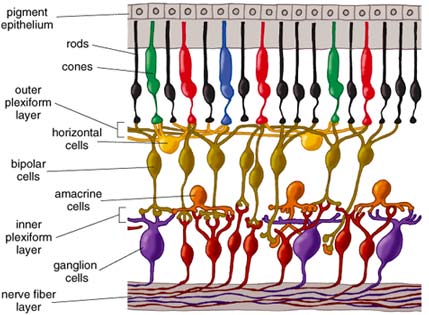
Understanding the anatomy of the primate retina is essential to understanding its function. Again, the photoreceptors lie in a layer against the back of the eyeball. In the second of three cell layers, called the inner nuclear layer, lie one to four types of horizontal cells, 11 types of bipolar cells and 22 to 30 types of amacrine cells. The numbers vary depending on species. The surface layer of the retina contains about 20 types of ganglion cells. Impulses from the ganglion cells travel to the brain via more than a million optic nerve fibers. The spaces separating these three layers are also anatomically distinct. The region containing synapses linking the photoreceptors with bipolar and horizontal cell dendrites is known as the outer plexiform layer; the area where the bipolar and amacrine cells connect to the ganglion cells is the inner plexiform layer.
Nicolas Cuenca and Helga Kolb
Decades of anatomical studies have shed light on how the retina works. Imaging techniques ranging from old-style Golgi silver staining, first used over a century ago by Ramón y Cajal, to electron microscopy and modern-day antibody staining have revealed the shapes and sizes of the retina's cell types and how the different cells connect to form synapses. Staining techniques have revealed electrical junctions between cells and the identity and location of neurotransmitter receptors and transporters. We now know that the neurotransmitter (chemical signal) passed through the vertical pathways of the retina—from photoreceptors to bipolar cells to ganglion cells—is glutamate. The horizontal and amacrine cells send signals using various excitatory and inhibitory amino acids, catecholamines, peptides and nitric oxide.
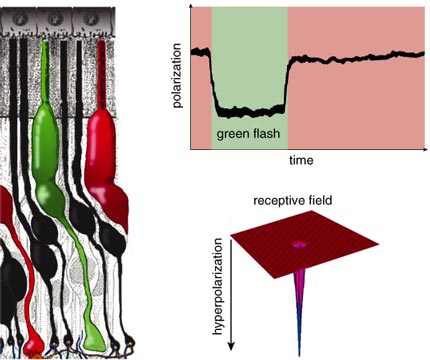
Helga Kolb
Electrophysiological investigations of the retina started 60 years ago. Studies of the optic nerve fibers showed that they could be stimulated to give traditional depolarizing action potentials, like those observed in other neurons. However, the first recordings of impulses within the retina by Gunnar Svaetichin in the 1950s showed very odd responses to light. Neurons in the outer retina—it was not immediately clear which cells he was recording from—responded to stimulation not with depolarizing spikes but with slow hyperpolarization. These "S potentials" are now known to originate with the photoreceptors and to be transmitted to horizontal cells and bipolar cells. The membrane hyperpolarization starts on exposure to light, follows the time course of a light flash and then returns to the baseline value when the light is off. This reflects the counterintuitive fact that both rods and cones release neurotransmitters during the dark, when the membrane is depolarized and sodium ions flow freely across the photoreceptors' cell membranes. When exposed to light, ion channels in the cell membranes close. The cells go into a hyperpolarized state for as long as the light continues to shine on them and do not release a neurotransmitter.
Although both rods and cones respond to light with a slow hyperpolarizing response, they report quite different image properties. Rods, detecting dim light, usually respond to relatively slow changes. Cones, dealing with bright signals, can detect rapid light fluctuations. In both cases, photoreceptors begin the process of decomposing images into separate parts. Both rods and cones respond to light directly over them. Thus, their receptive fields are very narrow.
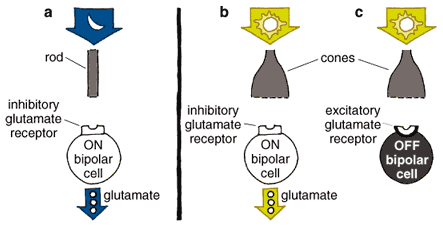
Tom Dunne
An image continues to be broken into component elements at the first synapses of the visual pathway, those between photoreceptors and bipolar cells. Different bipolar cells have different types of receptors for the neurotransmitter glutamate, allowing the cells to respond to photoreceptor input differently (Figure 7). Some bipolar cells are tuned to faster and some to slower fluctuations in the visual signal; some glutamate receptors resensitize rapidly and others more gradually. The cells thus fire either quickly in succession or relatively slowly in response to the same amount of stimulation. These receptors respond to glutamate by activating what's known as an OFF pathway in the visual process, detecting dark images against a lighter background. (Recall that photoreceptors constantly release glutamate unless exposed to light.) Other bipolar cells have inhibitory glutamate receptors; in other words, they prevent the bipolar cell from firing when the cell is exposed to the neurotransmitter. These receptors activate the ON pathway, detecting light images against a darker background.
The parallel sets of visual channels for ON (detecting light areas on dark backgrounds) and OFF (detecting dark areas on light backgrounds) qualities of an image are fundamental to our seeing. Vertebrate vision depends on perceiving the contrast between images and their backgrounds. For example, we read black letters against a white background using the OFF channels that start in the retina. Parallel bipolar channels transmit inputs to ganglion cells. Early in development the architecture of the inner plexiform layer, full of synapses between bipolar and ganglion cells, shows that synaptic connections become segregated in distinct, parallel pathways. Connections occur between ON bipolar cells and ON ganglion cells and also between OFF bipolar cells and OFF ganglion cells in demarcated portions of the inner plexiform layer.
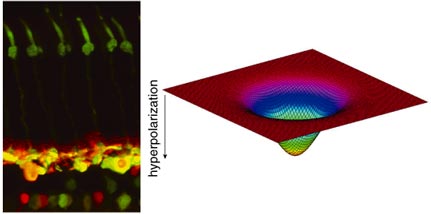
Nicolas Cuenca and Helga Kolb
If the retina were simply to transmit opposite-contrast images directly from the photoreceptors to the brain, the resulting vision would probably be coarse-grained and blurry. Further processing in the retina defines precise edges to images and allows us to focus on fine details. The honing of the image starts at the first synaptic level in the retina, where horizontal cells receive input from cones. Each horizontal cell actually receives input from many cones, so its collection area or receptive field is large. Horizontal cells' receptive fields become even broader because their plasma membranes fuse with those of neighboring horizontal cells at gap junctions. The membrane potentials of a whole sheet of cells become the same; consequently, horizontal cells respond to light over a very large area. Meanwhile, a single bipolar cell receives input from a handful of cones and thus has a medium-size receptive field.
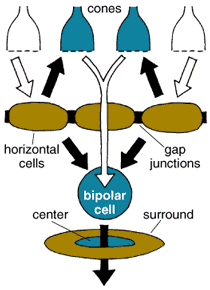
Tom Dunne
Whereas a single bipolar cell with its OFF or ON light response would carry a fairly blurry response to its ganglion cell, horizontal cells add an opponent signal that is spatially constrictive, giving the bipolar cell what is known as a center surround organization (Figure 9). The bipolar center signals either ON or OFF, and the horizontal cells add an OFF or ON surround signal, by one of two means. The horizontal cells can either signal the bipolar cell or feed information back to the cone photoreceptors themselves, which then feed forward information to the bipolar cells the cones contact. Feedback to the cones is now proposed to occur by means of an unusual electrical synapse consisting of half a gap junction; these hemi gap junctions are thought to change the ionic environment across the membrane of the cone photoreceptor. This complicated circuit from horizontal cell to cone to bipolar cells is still a subject of hot debate in the community of retina scientists.
Horizontal-cell function has occupied many vision scientists for decades, and much is now known about the role of these cells in the organization of visual messages. Horizontal cells respond to more than the photoreceptors that link to them. Feedback signals from the inner plexiform layer influence horizontal-cell activity as well. These feedback signals are transmitted via substances such as dopamine, nitric oxide and retinoic acid. The result is that horizontal cells modulate the photoreceptor signal under different lighting conditions—allowing signaling to become less sensitive in bright light and more sensitive in dim light—as well as shaping the receptive field of the bipolar cells, as we have seen. The horizontal cells can even make the bipolar cells' response color-coded, all apparently through feedback circuits to the cones.
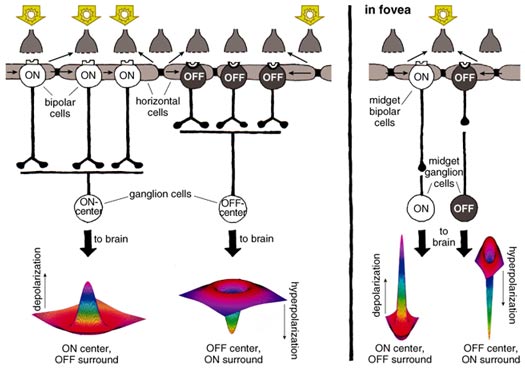
Tom Dunne
The ganglion cells have a receptive field organized as concentric circles. The amacrine-cell circuitry in the inner plexiform layer conveys additional information to the ganglion cell, possibly sharpening the boundary between center and surround even further than the horizontal-cell input does. In human retinas, two basic types of ganglion cells—ON center and OFF center—form the major output of the retina to the visual centers in the brain (Figure 10, left). ON-center ganglion cells are activated when a spot of light falls in the center of their receptive field and are inactivated when light falls on the field's periphery. OFF-center ganglion cells react in the opposite way: Their activity increases when the periphery of their receptive field is lit and decreases when light falls on the center of the field. (The receptive fields of ganglion cells are modeled as the difference between Gaussian distributions, giving them a so-called Mexican-hat shape.)
In contrast to the rest of the retina, the human fovea contains midget ganglion cells, which have minute dendritic trees connected in a one-to-one ratio with midget bipolar cells (Figure 10, right). The channel from midget bipolar to midget ganglion cell carries information from a single cone, thus relaying a point-to-point image from the fovea to the brain. Each red or green cone in the central fovea connects to two midget ganglion cells, so at all times each cone can either transmit a dark-on-light (OFF) signal or a light-on-dark (ON) message. The message that goes to the brain carries both spatial and spectral information of the finest resolution.
Nicolas Cuenca and Helga Kolb; Tom Dunne
Messages from blue cones are not processed in the same way as from red and green cones for some reason, possibly because the blue system is older in evolutionary terms. Blue cones are found in the retinas of most species. The typical mammalian retina also has green cones; primates have the additional red cones. Blue cones transmit information through a special blue cone bipolar cell to a different type of ganglion cell, which can carry both a blue ON and a yellow OFF response.
Electrical recordings show that several types of ganglion cells do not have concentric organization, especially in animals whose eyes lack a fovea. This includes most nonmammalian species and mammalian species that have retinas with visual streaks. Compared with species with foveas, the species with visual streaks do even more image processing in the retina itself before sending a message to the brain; their retinas can immediately synthesize information about image motion and direction of motion.
There is more to understand about the messages ganglion cells receive before they transmit a signal to the brain. For that, it is important to appreciate the organization of the inner plexiform layer, where 22 or more different types of amacrine cells make synaptic connections with about 20 different types of ganglion cells.
It was already clear from Cajal's description in the 19th century that amacrine-, ganglion- and bipolar-cell dendrites and axons were organized into distinct layers; Cajal himself divided the inner plexiform layer into five strata. But what sorts of synapses were formed among the tangle of intermeshing processes and what this organization meant were not immediately apparent. Electron microscopy helped to unravel this neurocircuitry. Now the interconnections of nine types of bipolar cells, 14 types of amacrine cells and eight types of ganglion cells are understood quite well. We can say we are half way to the goal of understanding the neural interplay between all the nerve cells in the retina.
Much is now known about what types of neurotransmitters different amacrine cells contain and about the organization of receptors at the different synapses. Amacrine cells are about equally divided between those that use glycine and those that use GABA (gamma-aminobutyric acid) neurotransmitters.
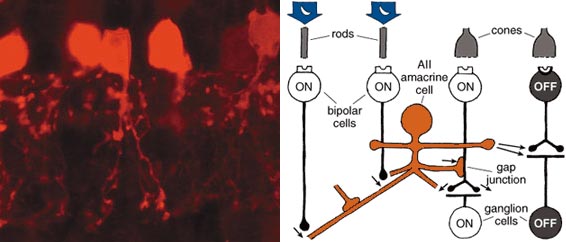
Nicolas Cuenca and Helga Kolb
Glycinergic amacrine cells are usually "small field." Their processes can spread vertically across several strata within the inner plexiform layer, but they extend relatively short distances horizontally. Glycinergic amacrine cells receive information from bipolar cells and transmit information to ganglion cells and to other bipolar and amacrine cells. Some glycinergic amacrine cells provide interconnections between ON and OFF systems of bipolar and ganglion cells. The most famous of these is called the AII cell; the AII and a GABA-releasing amacrine cell called A17 are pivotal in the circuitry of rod-based, dim-light vision in the mammalian retina. These cells aren't found in mammalian species that are active solely in daylight and have very few rods—for example, squirrels.
In the earlier discussion of ON and OFF channels emanating from cones, I neglected to talk about the channels from rod cells. Whereas cones connect in a direct pipeline to bipolar cells to ganglion cells, the bipolar cells that receive input from rods do not synapse with ganglion cells directly. The bipolar cells connected to rods are all of one type, solely transmitting an ON signal, and use the AII and A17 amacrine cells as intermediaries to get signals to ganglion cells. The small-field AII cell collects from about 30 rod-connected bipolar cells and transmits a depolarizing message both to ON (light-detecting) cone bipolar cells and to their ON ganglion cells and to OFF cone bipolar cells and OFF ganglion cells (Figure 11). It is as if the AII cells developed in the rod-dominated parts of the retina as an afterthought to the cone-to-ganglion cell architecture and now takes advantage of the preexisting cone pathway circuitry.
At the same time, the A17 amacrine cell collects rod messages from thousands of rod-connected bipolar cells. It somehow amplifies and modulates the information from the rod bipolar cells to transmit to the AII cells, but how it does this is not completely understood. In any case, the rod pathway with its series of convergent and then divergent intermediary neurons is clearly well designed to collect and amplify scattered vestiges of light for twilight and night vision.
Wide-field amacrine cells sometimes stretch horizontally across the inner plexiform layer for hundreds of microns and interact with hundreds of bipolar cells and many ganglion cells. Such amacrines are usually confined to one of the five different strata of the inner plexiform layer and create elegant meshworks of dendrites. Usually, they emit GABA as a neurotransmitter. Sometimes they connect to neighboring amacrine cells by gap junctions, increasing their sphere of influence and the speed at which signals transmit across large areas of retina.
Most GABA-releasing amacrine cells also release at least one other neuroactive substance. The secondary substances are usually neuromodulators rather than fast-acting neurotransmitters. The substances include peptides—"substance P," somatostatin, vasointestinal peptide and cholecystokinin—as well as the more familiar biomolecules serotonin, dopamine, acetylcholine, adenosine and nitric oxide. A variety of receptors have been found on ganglion and bipolar cells—for example, receptors for peptides, nicotine and muscarine (mushroom toxin) in addition to different forms of GABA receptors—indicating that amacrine cells are releasing such agents. Most of these neuromodulators are not active at conventional synapses; their release is thought to influence neurons even at a distance by diffusion. Such neuromodulators apparently influence the retinal circuitry under changing light conditions or even cause retinal activity to reflect the different times of day in the circadian clock.
A specialized amacrine cell releases dopamine when the retina is stimulated with intermittent flashing light. Dopamine causes the gap junctions among horizontal cells to become uncoupled, reducing the size of their receptive fields. Furthermore, the neurotransmitter affects the glutamate receptor on horizontal cells so that the amplitude of the light response declines. Again in the inner plexiform layer, dopamine closes gap junctions, this time the ones that link AII amacrine cells in large networks. The resulting uncoupling of the AII cells makes the effective field of influence of the rod-system amacrine cells much less significant in lighter conditions. Similarly in bright light conditions, another wide-field amacrine cell releases nitric oxide to uncouple the AII cell from the cone-bipolar system. All this removes the interference of the large-field rod pathway from the narrow-field cone pathways.
The above broad sketch of retinal circuitry suggests that the retina is remarkably complex. As vision research advances, the retina seems to take on an increasingly active role in perception. Although we do not fully understand the neural code that the ganglion-cell axons send as trains of spikes into the brain, we are coming close to understanding how ensembles of ganglion cells respond differently to aspects of the visual scene and how fields of influence on particular ganglion cells are constructed. Much of the construction of the visual images does seem to take place in the retina itself, although the final perception of sight is indisputably done in the brain.
Given how much is now known, it might be fair to ask, are we finished with the retina, or are there more surprises on the horizon? Earlier surprises included finding that much of the information transfer depended on electrical connections among cells rather than standard chemical synapses. For example, the major neural pathway from the rods depends on direct electrical connections. Some other fast-acting signals pass from amacrine cells into ganglion cells at gap junctions. Neuromodulators change the milieu of the neuron circuits but act from a distance by diffusion rather than at closely apposed synapses. Again, this is a surprising concept compared to the previous view that all neural interactions take place via neurotransmitters at specialized isolated patches of membrane apposition—that is, synapses. The most recent surprise has been that a previously unknown ganglion cell type appears to function as a giant photoreceptor itself, without needing input from rods or cones. This ganglion's cell membrane contains light-reactive molecules known as melanopsins. Given such unexpected findings, it appears that there may still be much more to learn about how the retina works.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.