How the Owl Tracks Its Prey
By Masakazu Konishi
Experiments with trained barn owls reveal how their acute sense of hearing enables them to catch prey in the dark.
Experiments with trained barn owls reveal how their acute sense of hearing enables them to catch prey in the dark.

DOI: 10.1511/2012.99.494
Editor’s Note: This Classic article was first published in the July–August 1973 issue, and is reprinted as part of American Scientist’s centennial year celebration. The author is recognized for his research on prey capture auditory systems in owls and singing in songbirds. Konishi received his B.S. and M.S. in zoology from Hokkaido University in Japan before coming to the University of California at Berkeley, where he obtained his Ph.D. in zoology in 1963. After two years in Germany in postdoctoral positions at the University of Tubingen and at one of the Max-Planck Institutes, he returned to the United States, working as a professor first at University of Wisconsin, then at Princeton University, and finally at California Institute of Technology, where he spent the rest of his career. He was elected into the National Academy of Sciences in 1985. Dr. Konishi’s current address: Division of Biology, MC 156-29, California Institute of Technology, Pasadena, CA 91125; konishim at caltech dot edu.

Payne and Drury (1958) were the first to demonstrate the ability of the barn owl (Tyto alba) to locate mice acoustically in total darkness. In a series of experiments, I have replicated their observation. A barn owl in pursuit of a mouse in the dark flies about 3.6–4.0 meters per second; it will fly faster if the mouse is visible or more slowly if the identity of the target is uncertain. As the owl comes within a range of about 60 centimeters from the mouse, it brings its feet forward and spreads the talons in an oval pattern. Just before hitting the mouse, it stretches its legs forward with the face and the wings lagging behind, often closing its eyes during this last phase of the strike.
The mouse does not seem to die instantly despite the powerful impact of the strike. Soon after landing, the owl always manages to bite the back of the mouse’s neck to kill it. Should the owl miss the mouse it will remain motionless on the ground and listen to the mouse in order to strike again from the landing site. If the owl can see the mouse hide behind the wall or under the floor, it will eagerly search for and run after it like a cat.

Photograph courtesy of M. Konishi
In the infrared photograph in Figure 1 it appears as if the owl were looking at the tethered mouse as it is about to strike. This worried me a little, since I could see through three layers of infrared filters the strobe filaments glow red as they fired. Of course, the owl could catch mice without the infrared strobes. My worry was whether or not the pictures I was taking depicted the true behavior of the owl in total darkness. In order to clear this doubt I repeated a clever experiment conducted by Payne (1962).

Photograph courtesy of M. Konishi
When a mouse walked quietly on foam rubber towing a rustling piece of paper several inches behind its tail, the owl tried to strike the paper instead of the mouse. Figure 2 shows the owl preparing to land on the paper, without noticing the mouse a small distance away. Besides demonstrating that the owl cannot see the mouse, this experiment proves two other important points: the owl cannot locate the mouse either by its smell or by its body heat (infrared radiation). (See Payne 1962 and 1971 for earlier papers on prey capture by owls with methods other than passive sound location.)
The above and later experiments might give the reader the impression that the owl strikes any sources of noise indiscriminately. Quite the contrary is true; the owl will not strike sounds new to it. Also, it can learn quickly slight differences between sounds bringing reward and no reward. If the owl has associated the appearances and sounds of prey and enemy a few times, it should be able to discriminate between the two in the dark by hearing alone.
M. Konishi
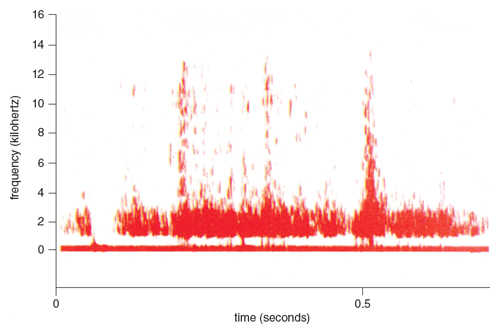
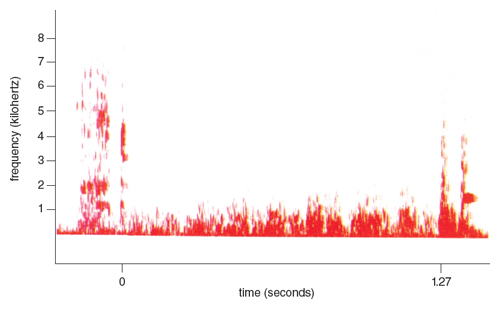
The rustling noises of the prey contain all the information needed for the owl to locate it in space. In order to design the later experiments, precise knowledge of the physical characteristics of these noises was needed. Since the vole is the main diet of the barn owl in the northeastern United States, I recorded and spectrographically analyzed the noises made by a vole moving through its subterranean hay-lined tunnel runways in a 20-gallon container within a soundproof room. The rustling noises contain a wide range of frequencies, as shown in Figure 3, but these data alone do not mean anything without knowing the auditory capability of the owl, and thus the hearing threshold of the barn owl had to be determined.
M. Konishi
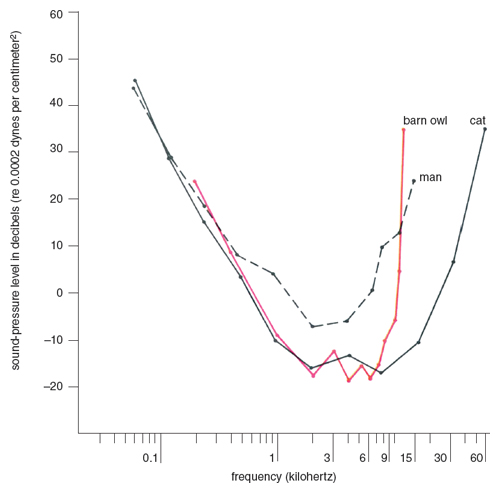
Three owls were used for this purpose. They were trained to take off from the perch for reward when they could hear a tone. Figure 4 presents the results from the owl that was most carefully tested. The owl’s hearing curve was drawn by connecting sound levels at which it responded correctly 75 percent of the time. The figure also compares the audibility curves of man, cat, and barn owl. Note that the cat and the owl have very similar auditory sensitivities up to about 7 kilohertz, beyond which the cat continues to be sensitive, while the owl’s sensitivity starts declining sharply. Both animals are much more sensitive than man in the frequency range from about 500 hertz to 10 kilohertz. No other birds that I studied are so sensitive as the barn owl, although some songbirds may be able to hear frequencies as high as the barn owl can (Konishi 1970).
Besides these quantitative data, I compared people and the owls under the same conditions. The owls could hear sounds which were so faint that none of my young undergraduate students and assistants could register them at the distance of the owl’s perch; however, above 12 kilohertz man is more sensitive than the barn owl. The barn owl thus can hear a large portion of the prey’s rustles, but it does not necessarily follow that the entire audible part of the rustles is equally important for sound location by the owl.
If the owl can be trained to locate electronically generated sounds, the cue effectiveness of various acoustic parameters can be analyzed under rigorously controlled conditions. I trained three owls to strike in the dark protected loudspeakers emitting various sounds of known physical properties. This I did in the following manner.
Since the hand-reared owls used in this work had had no experience catching mice in the dark, the training was done in several steps. The owls were first allowed to catch live mice under dim illumination and then in the dark. When pure tones and noises were broadcast through an earphone placed next to a dead mouse in dim light, the owls quickly learned to associate the artificial sounds and the mouse. After this step, they struck in the dark the earphone emitting those sounds.
Since the owls would stop striking the target as soon as they had eaten two mice, I constructed a device to dispense small pieces of meat as rewards for accurate location of the target. The owls learned to eat from the feeder within a few days. In the final stage of the training, the owls struck protected loudspeakers in the dark, then moved to the feeder for reward under dim illumination, and returned to the perch to wait for the next signal.
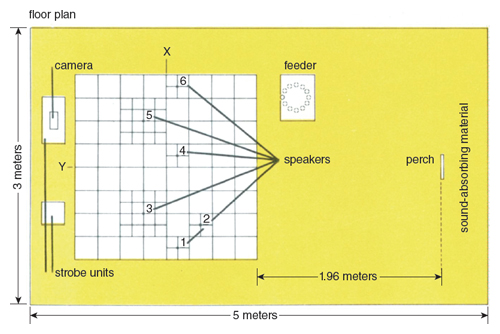
The accuracy of location was measured by an electronic device that registered the position of the owl as it struck the floor. It consisted of 100 square masonite plates, 10 centimeters x 10 centimeters and 20 centimeters x 20 centimeters, laid out like a chessboard covering the part of the floor where the owls were trained to land. The smaller plates surrounded the speakers, and the larger ones filled the remaining space. These plates were padded with foam rubber so that the owl could strike them hard without damaging the talons.
When the owl struck the plates, the microswitches installed beneath them closed and turned on small neon lamps which projected the chessboard pattern in a reduced size onto a panel. Six loudspeakers were installed under the chessboard floor. Short rubber tubing led sound from the loudspeakers to fixed intersections on the surface of the chessboard. The distance between the owl’s position and any one of the speakers could be read immediately on the lamp panel.
M. Konishi
The resolution of this measuring system was satisfactory for the purpose of the work. It was adjusted to the owl’s talon spread, which covered an area slightly larger than one 10 centimeters x 10 centimeters plate. When only one plate was struck, the midpoint between the owl’s feet was always close to the center of the plate. A slight deviation from the center would cause the owl to step on an adjoining plate, which means that little would be gained by using plates smaller than 10 centimeters x 10 centimeters. All training and tests were done in a soundproof, anechoic room 5 meters in length and 3 meters in width and height. The general layout of the room is shown in Figure 5. (Other technical details and the statistical treatments of the results are partly covered in Konishi 1973.)
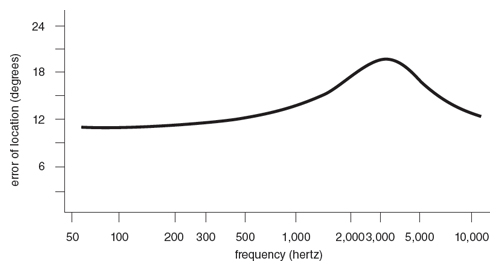
Let us first consider how man locates pure tones in order to provide some theoretical framework for experiments with owls. Man can locate pure tones by binaural comparison of intensity, phase and time of arrival. We discuss here the first two methods. Figure 6 shows the angular errors of location of pure tones of different frequencies in man. Notice that man can locate low and high frequencies rather well. There is a curious hump around 2–4 kilohertz where man makes larger errors. The theory to explain these results is as follows.
Tones of long wavelengths (i.e., low frequencies) bend around the head without creating intensity differences in the sound field around the head, whereas shorter wavelengths (higher frequencies) can be bounced back by the head, causing differences in sound intensity around the head. Since the magnitude of intensity differences between two ears varies with the direction of sound propagation, man can determine the direction by binaural comparison of intensity. The shorter the wavelength relative to the diameter of the head, the more distinct is the sound shadow so created. Therefore, man can locate higher frequency tones relatively accurately.
Low-frequency tones are located by detecting phase differences between the ears, which are due to differences in the paths traveled by sound to reach the two ears. For each frequency, the magnitude and sign of phase differences vary according to the position of the sound source relative to the median plane of the head. This is the basis for location by binaural phase comparison.
For this method to be effective tones of wavelengths longer than at least twice the interaural distance are necessary, that is, d < λ/2, where d is the distance between the ears and l wavelength. When d > λ/2, a binaural phase difference of more than 180 degrees results, and it be comes impossible to discern which ear is in the leading phase, since a phase difference of 180° + φ is equivalent to an opposite phase difference of 180° - φ (from 180° + φ - [180° - φ] = 360° = 0, where φ is the excess angle over 180°). It is this ambiguity that makes the phase method ineffective with higher frequencies (Gulick 1971; Mills 1972; Steven and Newman 1934).
After Steven and Newman 1934.
These conditions, higher frequencies for intensity comparison and lower frequencies for phase comparison, create for man a frequency range (2–4 kilohertz) in which neither the phase nor the intensity method is very effective. This explains the hump in Figure 6. Whether or not the above theory applies to the owl, it can suggest useful research strategies.
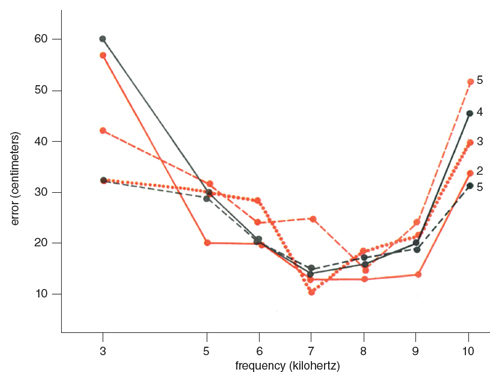
Since continuous pure tones can produce differences between the ears only in two acoustic parameters, namely intensity and phase, they are suitable for analyzing the acoustic method used by the owl. Tone signals were broadcast at a constant intensity of 4 decibels (re 0.0002 dynes per centimeter squared) at the perch. The signals lasted until the owl landed. The results from one owl are graphically summarized in Figure 7. Low- and high-frequency tones such as 3 kilohertz and 10 kilohertz were harder for the owl to locate than those between 6 and 9 kilohertz. These differences in the error of location are due neither to the variation in the owl’s auditory sensitivity nor to the directionality of the speakers, both of which depend on frequency. Adjustment in sound intensity according to the owl’s audibility curve did not significantly affect the error curves. The speakers did not become sharply directional at higher frequencies.
M. Konishi
The results can be partly explained in terms of binaural intensity comparison, although this cannot account for the sudden increase in the error of location above 10 kilohertz. The barn owl does not seem to use the phase method, at least in the same way that man does, because it located low-frequency tones poorly and because it did not have any intermediate frequency range in which the error of location increased. Since the distance between the owl’s ears is shorter than that of man, the frequency range unsuitable for both the intensity and phase methods, if it exists, should be higher for the owl than for man.
The mouse rustles are not steady but discontinuous noises. Abrupt inflections in these noises would be useful for binaural comparison of time. Instead of binaural phase differences of a continuous tone, the time method uses differences in time of arrival which are caused by differences in the paths traveled by the first wave of sound. This method is independent of frequency. A series of tone beeps should provide the owl with sufficient time cues, because each beep has an onset and a cutoff. I compared the errors of location obtained by using tone beeps (50 or 100 milliseconds in duration separated by silent intervals of 80 or 150 milliseconds) and sustained tones and found no consistent differences between them. Moreover, just as the errors of location with sustained tones depended on frequency, so did those with tone beeps (Figure 7).
In theory, a single pure tone should be hard for the owl to locate, because even two ears are insufficient to define a point in three dimensions. For example, there is not one but a family of points around the head where a tone can produce a given inequality in intensity between the ears. The relatively good location of some pure tones by the owls is perhaps due to head movements. The rustling noises of prey contain many frequencies, as mentioned earlier, and the reason why the owl can use them so effectively will be sought next. The results of the pure-tone tests should provide bases for useful predictions. It would also be possible to find out which components of the noises are used by the owl by systematically removing different parts. However, I decided to use a more systematic and controllable approach.
I constructed signals with a constant center frequency and different bandwidths (maximum frequency minus minimum frequency) to study the effects of bandwidths on the error of location. The signals were broadcast at a constant intensity of -12 decibels (re 0.0002 dynes per centimeter squared) at the perch and lasted until landing. For comparison with the noise signals, the errors of location were measured for a 7 kilohertz tone delivered at an intensity of 4 decibels (re 0.0002 dynes per centimeter squared). The noise signals were delivered at a lower intensity in order to demonstrate their superiority as location cues over the most effective puretone broadcast at a higher sound level.

Since the differences in the errors of location were small, they were compared by scoring and statistical procedures that were immune to the errors in estimating the owl’s position. The distribution of strikes in Table 1 shows a general tendency for larger proportions of strikes to fall within the four-plate zone and its immediate vicinity with increasing bandwidths. Noises containing frequencies between 5.5 and 9.5 kilohertz were more accurately located than those involving other frequency ranges. A 4 kilohertz band noise centered around 7.5 kilohertz is sufficient for accurate location. Additional frequencies do not contribute to more accurate location.
The owl needs, therefore, only a small portion of the frequency spectrum in the prey’s rustles. No wonder the owl can precisely locate small rodents that make wideband noises rich in frequencies in the range most suitable for sound location. It should be noted that 5.5–9.5 kilohertz is the range in which the barn owl is most sensitive, although a higher sensitivity itself does not contribute to more accurate location, as mentioned before.
Small rodents make noises by moving. How does the barn owl catch a moving prey in the dark? What happens if, after the owl takes off, the prey moves or stops making sound? When I simulated these conditions electronically, I found that all three owls made larger errors when the signal stopped upon take-off than when it continued until landing (Figure 8). For example, when the signal continued until landing, one owl hit 46 times out of 58 trials within the area covered by the four 10 centimeter x 10 centimeter plates surrounding the target, while the same bird missed that area 69 times out of 86 trials in the absence of a post-take-off signal. Making the signal louder did not help the owl locate the target better without post-take-off signals.
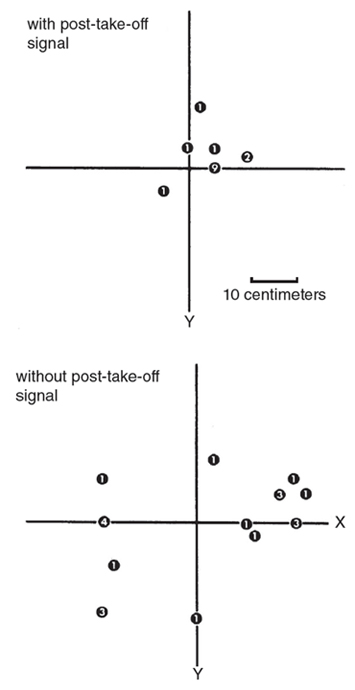
M. Konishi
The above results suggest that the owl can make mid-flight course corrections, like the moon shots, in order to strike the target accurately. Small rodents make noises intermittently, and the owl must be able to adjust to this condition. In another series of tests, I let the signal stop upon take-off and reappear after the owl had flown for varying periods. When the signal reappeared after the owl had flown for 0.5 second out of the total flight time of 1.2 seconds, the owl still struck the target as accurately as when the signal continued until landing.
The accuracy of location was not affected until about 80 percent of the total flight time was devoid of signal. When the owl had to fly for a period of 1 second without signal, it located the target as poorly as when the signal stopped completely upon take-off. Another factor that affects mid-flight corrections is the timing of post-take-off signals. When the owl could hear a faint and brief (50 milliseconds) noise burst three times (0.3, 0.6, and 0.9 second after take-off), it could locate the target as accurately as with a continuous noise.
The most crucial test of the owl’s ability to make mid-flight corrections involves the use of two loudspeakers: the signal shifts from one speaker to the other during flight. Figure 9 shows an owl changing its flight direction as the signal shifted from one speaker to another. Notice the direction of the owl’s face. It turns its face toward the new target position before orienting its body.

Photograph courtesy of M. Konishi
In Figure 9 infrared flashes were delivered at constant intervals of 250 milliseconds. Notice that the second and third exposures are closer together than the others; this is because the owl reduces its flight speed as soon as it hears a shift in the target position. When the owl has to make a large course correction, it comes to a sudden halt in midair and hovers before advancing toward the new target position. Because of this deceleration and the longer flight path required, the owls took a significantly longer time to reach a speaker when it was used as a second target than as a single source.
To hear faint and brief noises in flight and correct the flight course must be a difficult feat. One would wonder how the owl manages to do this when its own wing noises might mask the signal. Owls are known to fly much more quietly than other birds. Their body feathers are soft, and the leading edge of their wings has a fine comb, which is supposed to suppress the wing noises (Graham 1934). A recent study, however, reports that the removal of the comb had no effect on the wing noises of the tawny owl (Neuhaus, Bratting, and Schweizer 1972).
When I recorded and analyzed the wing noises of one of the barn owls during location tests (Figure 10), I found that the flight noises are not only faint but also lack high-frequency components. Most of their energy is concentrated below 1 kilohertz; above 3 kilohertz there is too little energy to record even with a very sensitive set of equipment. Similar results were reported for other species of owls (Gruschka, Borchers, and Coble 1971; Neuhaus, Bratting, and Schweizer 1972).
M. Konishi
These findings imply that the owl’s wing noises would not interfere with the detection of acoustic clues for mid-flight correction, since useful cues are noises between 6 and 9 kilohertz. The lack of high frequencies is also advantageous for the owl, because small rodents capable of hearing high frequencies cannot hear and locate the approaching owl. The house mouse and some deer mice are rather insensitive to frequencies below 3 kilohertz (Ralls 1967).
Some rodents, such as the kangaroo rat, however, are quite sensitive to low frequencies, which might enable them to hear and discover the owl. The resonance frequency of the kangaroo rat’s middle-ear cavity, which is low due to its enlarged mastoid bulla, increases the sensitivity of the rat ear to low frequencies (Webster 1972). If the bulla cavities of a kangaroo rat are obliterated, its chance of being caught by an owl greatly increases, which is perhaps due to the inability of the rat to hear the flight noises of the owl (Webster 1962).
In some species, such as the barn owl and the saw-whet owl, the left and right ear openings differ from each other in their size and/or position. In the barn owl the ear openings are about the same in size, but the left one, together with the skin flap in front of it, is located higher than the right ear hole and skin flap (Figure 11).

Photograph courtesy of M. Konishi
There does not seem to be any individual difference in this asymmetry—no left- or right-handed owls. Also, in the barn owl the asymmetry is restricted to the ear opening without affecting the middle and inner ears. The binaural methods of sound location make use of the lateral displacement between the ears to determine the azimuth of the source. By the same token, the vertical displacement has been thought to enable the owl to determine the elevation of acoustic targets (Norberg 1968; Payne 1962, 1971).
I tested this idea by a simple experiment that involved plugging one ear. The owls (I used two) with one ear plugged veered toward the side of the target opposite to the plugged ear. This would be expected if the owl uses binaural comparison of intensity; the signal should sound louder to the intact ear, and thus the owl estimates the target position too far toward that side.
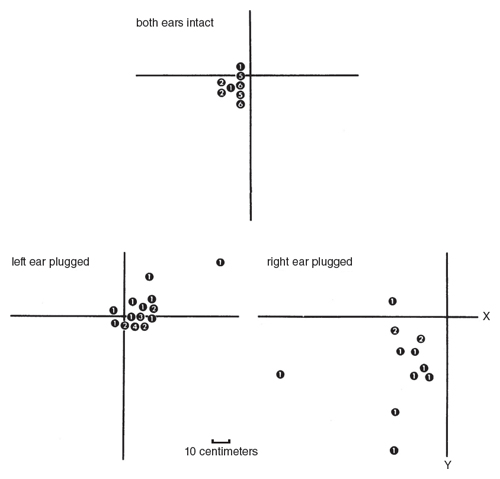
M. Konishi
One of the owls that I tested more extensively made systematic errors in the vertical direction. When its right ear was plugged, it struck short of the target, and blocking its left ear caused it to land slightly beyond the target on the average (Figure 12). These results suggest that the vertical displacement between the ears is not used in the same simple way described for the lateral displacement. To make the matter more complex, both owls struck closer to the target with their right ear unplugged than with their left ear unplugged. This may be due to differences in the degree of ear blockage, which could not be precisely controlled, although the same bird produced a similar set of results twice. It is also possible that the right ear plays a more important role than the left one in location. A person deaf in one ear is known to be able to locate sound. The pinna seems to play a crucial role in monoaural sound location in man (Batteau 1967).

Photograph courtesy of M. Konishi.
Owls do not have a structure homologous to the mammalian pinna, but some of them have a fold of skin extending from the forehead above the eye and along the orbit behind the ear to the base of the lower mandible. In the saw-whet owl, this skin fold is quite large around the ear. In the barn owl, the skin fold itself is not so prominent, but it carries a tall curved wall of densely packed feathers which encircles each half of the face. The left and right halves meet along the midline of the face, where the feather walls from the two sides form a pointed ridge (see Figure 13). When the owl is not attentive, this ridge broadens.
On each side of the face, the curved wall looks like a trough with a paraboloid inner surface. At the level of the ear opening, the skin flap covers the trough, forming a tunnel in which the ear hole is located. The entire facial structure makes up the well-known heart-shaped outline of the owl’s face, called the facial disc. When one sees the whole design of the facial disc, one cannot help thinking of a sound-collecting device.
Does the facial disc facilitate directional hearing? Payne (1962, 1971), using a stuffed barn owl, and Norberg (1968), using a stuffed Tengmalm’s owl, measured the directionality of the ears by monitoring sound near the eardrum. According to Payne, for low frequencies the barn owl’s ear is only moderately directional. For high frequencies above 8.5 kilohertz, the ear becomes highly directional and also the pattern of directionality reflects the vertical displacement of the ears. Slight changes in the shape of the facial disc and the orientation of the skin flap affected the pattern of directionality for higher frequencies.
I removed the facial-disc feathers of a barn owl to find out whether and how its errors of location would be affected. The owl was tested with a continuous noise broadcast at the lowest sound level that assured accurate location, so as to be able to detect any slight change in the accuracy of location in the absence of the facial disc.
The operated owl made large errors by landing short of the target. However, when I increased the sound level by 10 decibels, the owl improved its accuracy of location considerably. No greater improvement resulted with an increase of 20 decibels. A 5-decibel increase did not reduce errors at all (Figure 14). These observations suggest that the facial disc may be a sound amplifier; it collects sound from a large area and focuses it onto a smaller area. Payne (1962) thought the facial disc was too small to be an effective amplifier in the frequency range audible to the owl.
The amount of amplification (gain) of a paraboloid antenna is a function of its diameter and the wavelength of sound expressed as G = η(πD)/λ)2, where G is gain, η is the aperture efficiency and is larger than 0 and smaller than 1, D is diameter, and λ wavelength. The widest part of the facial disc is about 7 centimeters in diameter, and the wavelength of 7 kilohertz is 4.9 centimeters. Using these values in the above equation and assuming η = 0.5, we obtain G = 10, which means a gain of 10 decibels. This is a small amount of amplification but should be useful when the owl must detect faint noises.
The facial disc may not function as a paraboloid antenna, but the above calculation should provide some idea as to the operating conditions and effectiveness of such a sound-collecting device. The facial disc seems also to contribute to directional hearing, since the owl, even with increased sound intensities, failed to recover the degree of accuracy attained before the operation. The directionality of a parabola is also a function of its diameter, shape, and wavelength. It is not yet known to what extent the owl controls the shape of the facial disc and the orientation of the skin flap during sound location. Solution of these problems seems essential for the understanding of the mechanism of sound location in this species.
Payne (1962, 1971) used his directionality data to conclude that, if the owl moves its head so that the amplitudes of all frequencies are maximized at both ears, it must be directly facing the target. Since the ear becomes sharply directional for higher frequencies, these would help obtain a fine azimuthal bearing. Since the asymmetry of the ears causes a vertical displacement in their directionality at higher frequencies, these would enable the owl to align its head precisely in the vertical direction.
Two lines of evidence make this theory untenable. First, the barn owl does not need such high frequencies as 8.5–13 kilohertz, which Payne’s theory requires. The owl can locate noises containing frequencies between 6 and 8.5 kilohertz accurately. Second, a simple experiment will show that the theory fails to explain the ability of the owl to recognize the direction of sound before it moves its head. Man can locate sound quite well without head movement, although it seems essential in the absence of the pinna (Freedman and Fisher 1968). I have not tried to restrain the owl’s head, but I used a trick to get the same effect.
If the owl turned its head toward a signal lasting shorter than the time required to initiate or complete the turning of the head, the owl should not have been able to align its head direction with the target by successive steps of readjustment, which Payne’s theory requires. I examined the direction of the owl’s face in infrared pictures taken during tests in which the owl was allowed to hear only one brief noise burst to redirect its flight course from one speaker to another. In every case the owl’s head continued to turn well after the signal had stopped. The owl oriented its head in the general direction of a signal lasting as short a time as 10 milliseconds, which is too brief an interval for the owl to initiate head movement.
Pumphrey (1948) developed a theory for owls with asymmetric ears to explain the location of sound without head movement. This theory also uses the frequency-dependent asymmetry of the ears’ directionality. It requires two ears and at least three bands of frequencies. As mentioned before, there are many points around the head at which a tone can produce a given inequality in intensity between the ears. These points are contained in a surface; each band of frequencies defines a surface. Because of the asymmetry, some surfaces intersect one another, and three of them can define a point in space unambiguously. The test of this theory must be done without head movement.
There are other general theories of sound location which will not be described here. The errors of sound location discussed so far consist of two components—errors in auditory location and deviations in the control of flight direction. We have recently designed a different type of experiment to measure the true accuracy of auditory location without flight. This work is still in progress. My studies demonstrate what the barn owl can do under the experimental conditions used. In nature, it must hunt under different and varying conditions which might render some of these potentials unusable or require capabilities not uncovered by my studies. Combinations of field and laboratory experiments will be necessary to learn more about the natural acoustic behavior of the barn owl. It should also be emphasized that other species of owls may have different acoustic capabilities.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.