How Animals Communicate Via Pheromones
By Tristram Wyatt
Human behaviors are probably influenced by invisible smell signals, just like all other animals.
Human behaviors are probably influenced by invisible smell signals, just like all other animals.

DOI: 10.1511/2015.113.114
Powerful messages can be delivered by smell, a fact known long before the means by which they did so was discovered. The ancient Greeks knew that secretions from a female dog in heat were attractive to male dogs. In his beekeepers’ manual The Feminine Monarchie published in 1623, Charles Butler warned that an injured bee’s "ranke smell" would attract other angry bees to sting. In the late 19th century, New York’s first state entomologist, Joseph Lintner, noted the crowds gathered on the sidewalk outside, looking at the spectacle of 50 wild silk moth males attracted to a virgin female moth in his office window.

Photograph by Alexander Wild.
Lintner correctly observed that the attraction was probably a chemical, detected by the large, elaborate antennae of the males. He predicted that such an irresistible and far-reaching force could be used to control agricultural pest moths, if only chemists could identify and synthesize these powerful molecules.
Lintner’s prediction has been generously fulfilled (see below for "Putting Pheromones to Use”). But because the quantity of pheromone produced by each animal is so small, it was almost 70 years before the first pheromone could be chemically identified, in 1959, by the Nobel Prize–winning German chemist Adolf Butenandt and his large team.
In the half century since then, as the technology for isolating and identifying trace amounts of compounds has become more refined, pheromones have been found in almost every kind of animal, in squid, lobsters, ants, fish, salamanders, and mice, to name just a few. Although pheromones are important in many species for finding mates for sex, they can have a wide variety of other functions, such as one produced by mother rabbits that prompts suckling by their pups. In many social insects, such as the ants, bees, and wasps, almost every part of colony behavior is mediated by pheromones, from queen signals affecting worker reproduction within the colony to Butler’s "ranke smell" alarm pheromone that activates colony defense against enemies.
Pheromones are central to the lives of animals, and in my 30 years of studying pheromone evolution, I have come to appreciate the enormous variety of ways that animals use pheromones as well as how similarly these signals are perceived by smell. Studies of moths, social insects, and mice have contributed some of the biggest breakthroughs in pheromone research, but one member of the animal kingdom remains a tantalizing mystery: humans.
Pheromones are chemical signals that have evolved for communication between members of the same species. A pheromone signal elicits a specific reaction in the receiver, for example, a stereotyped behavior (releaser effect) or a developmental process (primer effect). Some pheromones can have both effects. All sorts of molecules, large and small, have been found acting as pheromones, depending on whether the message is sent out on wind or water currents or placed directly on the nose or antenna of the recipient.
Most pheromones are detected by the sense of smell. However, not all smells are pheromones. Mammals, including humans, also give off a cloud of molecules that represent our unique individual "smell" or chemical profile. These differences between individuals make it possible for dogs to distinguish people by smell. People are quite good at it, too—parents can distinguish their baby from others by smell alone. Ants similarly can distinguish between members of their own colony and those from other colonies. In both mammals and insects, learning is necessary to develop this ability. The sources of the molecules that make up an animal’s "individual smell" include its own secretions and also may reflect its environment, food, bacteria, immune system, and molecules picked up from other individuals in its social group.

Photograph courtesy of Sam Woo.
To definitively demonstrate that a pheromone exists, one must design a repeatable experiment, a bioassay, that shows that a smell molecule (odorant) causes a particular effect on the receiver, for example inducing a behavior. Butenandt used such a bioassay to enable his identification of the first pheromone in the silk moth: He observed that males showed excited wing fluttering in response to the smell of mature females (and extracts of their pheromone glands). The wing fluttering response gave Butenandt a bioassay to see which fractions of the extract contained the pheromone activity as he attempted the next steps of isolating the possible pheromone molecule. He and his team tested many extracts and identified the sought-after odorant when it induced the telltale fluttering behavior. They then synthesized the molecule and showed that the synthetic molecules had the same effect as real female extracts.
These steps are basically the same ones that need to be carried out today to identify the pheromones of any organism, starting with finding a good bioassay. What has changed is that, whereas Butenandt needed 500,000 female moths to get enough material for his analysis, a team today could attempt much of the analysis using just a few individual moths. This is largely due to improvements in technology, notably gas chromatography linked to mass spectroscopy, but also because we now often know the kind of molecules we are looking for.
Pheromones are smells that have evolved functions as signals. How the sense of smell works was a mystery until 1991, when Linda Buck and Richard Axel, then both at Columbia University, published their discovery of the highly variable family of almost 1,000 different olfactory receptors in the nose of the mouse. They received the 2004 Nobel Prize in Physiology or Medicine for this work, which opened a whole new avenue of pheromone research as the receptors, including those for pheromones, could now be identified.
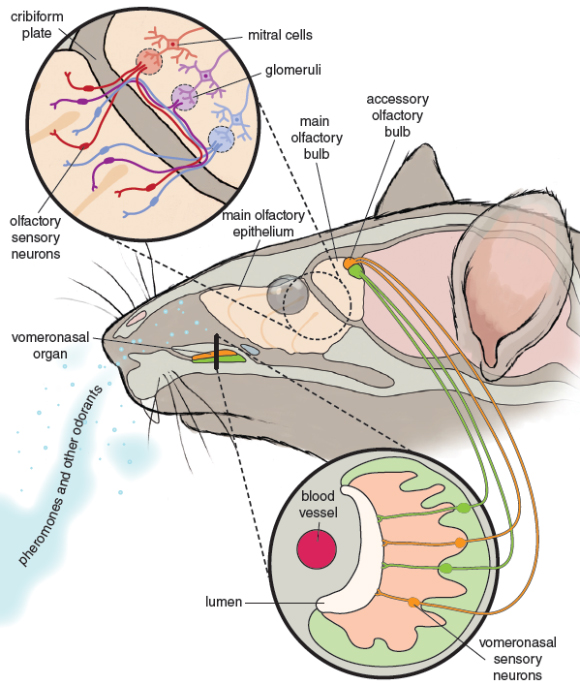
Illustration by Barbara Aulicino.
Odorant molecules interact with these chemosensory receptors, proteins on the surface membrane of olfactory sensory neurons exposed in the nose to the outside world. When an odorant binds to the chemosensory receptor, it stimulates the nerve to send a signal to the brain. Buck and Axel also discovered that each olfactory sensory neuron in the mouse nose carries just one of the 1,000 types of chemosensory receptor. All olfactory sensory neurons with the same receptor type go to the same collection point, a glomerulus in the olfactory bulb of the brain. There is a glomerulus for each receptor type.
Insect chemosensory receptors are so different from mammal ones that it took almost another decade to find them, and it is likely that they evolved independently from those in mammals.However, although the receptors in insects and mammals are very different, the "wiring diagram" organization of olfaction is remarkably similar, with all olfactory sensory neurons carrying the same type of receptor going to the same glomerulus in the brain. It could be that this organization evolved independently by convergence, because this is the best way of organizing chemosensory reception, or it may be that the similar organization is a relic of the way it was arranged in a distant shared common ancestor.
Small differences in the amino acid sequences of these chemosensory receptor proteins determine which odorant molecules bind to them. Most receptors are “broadly tuned”; that is, they are stimulated by a range of similar odor molecules. Conversely, most odorant molecules will stimulate a range of receptors. The combination of receptors stimulated by an odorant tells the brain what molecule or molecules have been smelled. This combinatorial code, together with the broad range of receptors, gives olfaction a great ability to distinguish different odorants.
The variety of molecules that animals have evolved to use as pheromones is enormous. The variety is explained by the ways that relevant molecules become co-opted as pheromones through evolution, my research focus. At a time when most scientists have had to become more narrowly focused, I have had the good fortune to have the opportunity to bring together the results of research on pheromones across all kinds of animals to create a unifying evolutionary synthesis, integrating behavioral and molecular levels.
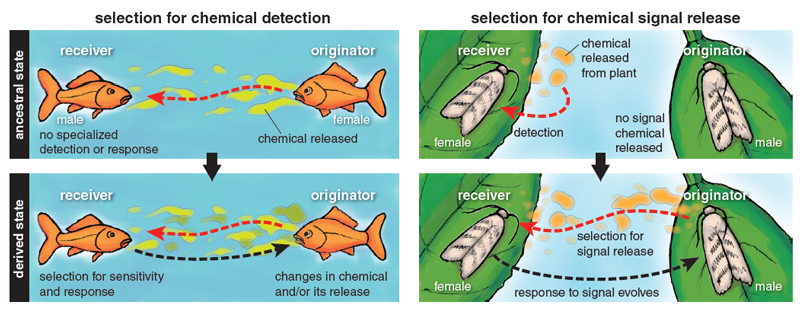
Illustration by Barbara Aulicino.
Odorant molecules became signal molecules in two main ways. The first way is by developing increasing sensitivity for molecules that initially are simply a reliable cue, for example, to finding a sexually mature female. This may be how goldfish sex pheromones evolved. Hormones control the development of eggs in female goldfish, and some of the hormones leak out of their gills into the water. These hormones, or molecules derived from them, have become female-produced sex pheromones, attracting male goldfish. Back in evolutionary time, some males could detect the hormones in the water, using their general broadly tuned olfactory receptors. The most sensitive males would find the females first, leading to selection over generations for greater sensitivity and increasing specificity of these receptors to avoid false alarms. There would in turn be selection on females to produce these hormone molecules specifically for release, because females benefit from attracting males to fertilize their eggs, leading to an evolved system of olfactory communication between a signaler and receivers.
The second way is by exploiting preexisting sensitivities of the receiver. The male sex pheromone of the European beewolf wasp Philanthus triangulum, (Z)-11-eicosen-1-ol, may have evolved this way. The male sex pheromone exploits a preexisting female sensitivity to this molecule, which is given off by the female beewolf’s prey, honeybees.
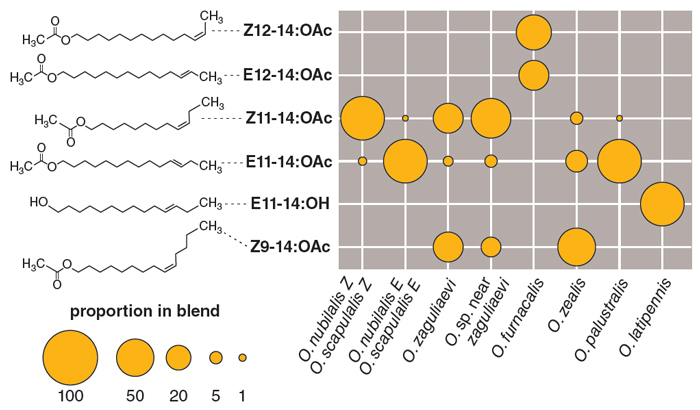
Adapted from J.-M. Lassance, et al., Proceedings of the National Academy of Sciences of the U.S.A. 110:3967–3972
In the literature on pheromones, much emphasis is given to their specificity, and their diversity is central to the story of species divergence and evolution. Most pheromones are not a single molecule: Instead, particular combinations of molecules form a multicomponent pheromone. When molecules are added or lost from the
cocktail of molecules as species diverge, each new species will have its own specific sex pheromone from these new combinations, even though the molecules themselves may not be especially unusual. For animals that use peptides or proteins as their pheromones, such as newts, a different amino acid sequence is used to gain specificity and avoid courting an animal from the wrong species.
Releasing pheromones can attract more than a mate. Just as a singing bird can be heard by and attract a cat, broadcast pheromones can be detected by some predators, and even more sinister, some predators can counterfeit pheromones to lure unsuspecting prey (see below for "Spying and Deceit").
Because many moth species are crop pests, the pheromones of this group have been particularly well studied for their potential in biocontrol methods. Moths provide model systems to explore how animals find their way to a pheromone source to find a mate and how animals distinguish between the pheromones of different related species.
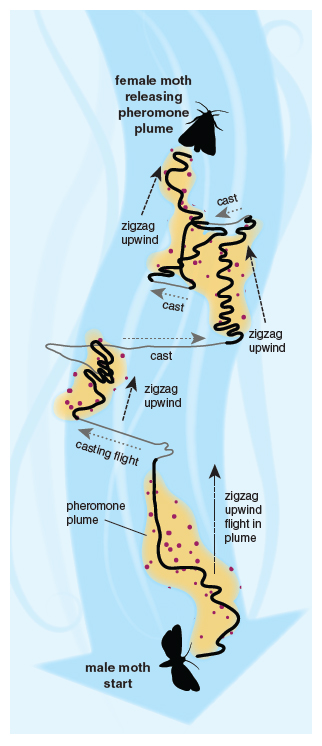
Illustration by Barbara Aulicino.
When a female moth releases her volatile sex pheromone into the night air, it is carried by the wind like an invisible smoke plume from a chimney. The wind shears it into pockets of air with pheromone separated by clean air pockets. When a pocket of air with pheromone hits the antennae of a male searching on the wing, he responds by flying upwind for a fraction of a second. If he hits another pocket of pheromone, he flies upwind again. If he doesn’t hit more pheromone, he flies from side to side until he hits pheromone again. This is a neat way of effectively flying upwind towards the female despite rapidly changing wind directions. This strategy has inspired programming of robots to orient up a chemical plume, and vertebrates such as fish and albatross show similar zig-zag tracks toward an odor source.

Photograph by Ann Pettigrew.
Most female moth sex pheromones consist of a specific combination of molecules. Closely related species use the same molecules in different ratios or share many, but not all, of the molecules: The differences allow males to distinguish females of their own species from those of other species. As the male flies upwind he will only respond if the pheromone hitting his antennae contains just the right combination for his species. On his antennae are the olfactory sensory neurons with specialized receptors. Moth males in species with multicomponent pheromones have one receptor specific for each molecule that makes up the pheromone. If all the right molecules are present, in the right ratio, then the right combination of nerve cells and their glomeruli are stimulated, sending a message to the higher brain: "Fly upwind." However, the male moths are also sensitive to distinctive molecules in the pheromone blends of related species. If a male moth detects those, he stops flying upwind to avoid wasting his time pursuing a female from another species.
Social insects have provided some of the most spectacular examples of pheromone evolution. Pheromones mediate many of the complex interplays within colonies of social insects such as bees, wasps, and ants. These include the familiar trails of ants and the honeybee alarm pheromones mentioned earlier. However, pheromones are also essential mediators of two aspects of the defining feature of social insects, the division of colony members into queens and workers. Only the queen (or a small number of queens) produces fertile eggs.
First, a pheromone "tells" the workers not to reproduce. The signal is a queen pheromone, worn like a crown only by the queen in an ant or honeybee colony. It marks her and also seems to tell the workers that because she is fertile and laying eggs, the workers do not need to do so. With colonies of up to half a million individuals, pheromones are the only practical way for the queen to send a message to all the workers in the colony.
In many species the workers’ ovaries do not develop if the workers detect the queen pheromone. If the queen is removed and her pheromone is no longer detected, then the workers start to develop ovaries. This property can be used to sensitively test which molecules produced by a queen can act in her absence. University of Leuven’s Annette Van Oystaeyen and her colleagues used this bioassay to explore queen pheromones in a wide range of ants, bees, and wasps in a recent international study. They showed that in most kinds of ants, bees, and wasps the chemicals are similar nonvolatile saturated long-chain hydrocarbons. These queen pheromones, which signal her fertility, are chemically similar to and may have evolved from sex pheromones advertising fertility in solitary ancestor species.
The second key role for pheromones in social insects is controlling which larvae become queens in the first place. This is a developmental (primer) effect of pheromones on the larvae. In the honeybee, any female larva has the genetic potential to develop into a worker or a queen, depending on what it is fed as a larva. If it is fed only royal jelly, a special food produced by nurse worker bees, the larva will develop irreversibly into a queen. The switch is triggered by the primer effects of pheromones in the royal jelly, including a protein called royalactin, recently identified by Masaki Kamakura of Toyama Prefectural University, Japan. Among the many effects of royalactin in the larva are changes in hormone release, which in turn may affect which genes are turned on and off via DNA methylation. The queen will be much larger than workers, live 20 times as long, and have functioning ovaries to lay up to a million eggs.
Primer effects also occur in mammals. Mice living together may affect each other’s reproductive development via primer effects. For example, exposure to the urine of dominant adult male mice causes immature female mice to become sexually mature more quickly. However, it is not clear which molecules in the male urine are inducing this primer effect. There have also been questions about whether these effects occur in the wild, because most have only been observed in the laboratory.
Vertebrates, including mammals, use pheromones extensively. In mammals, both small molecules and large proteins have been identified as pheromones. The range of mammals shown to use them includes elephants, goats, and pigs, but the best studied is the house mouse, building on its status as a key model species in medical research.
When we switch off the lights and go to bed, the house mouse is in its element. Like many nocturnal animals, it has large, sensitive eyes and ears. But much more important is its world of smells. A dominant male spends much of his time crisscrossing his territory, painting drops of his urine on landmarks, boasting his ownership. The volatile smells wafting from the urine marks attract females to come close to the scent mark, where they sniff up a nonvolatile protein pheromone, darcin. A message sent to the female’s brain prompts her to remember the individual smell of the male who left the mark and also remember its location. She will return to the spot and mate with the territory owner, recognized by his individual smell.
All vertebrates have a main olfactory system, the nose. Frogs, salamanders, snakes, and many mammals—but not humans and other higher primates—also have a second nose, the vomeronasal olfactorysystem. For a while it was thought in error that vertebrate pheromones were exclusively perceived via the vomeronasal system. It is now clear that vertebrate pheromones can be detected by either or both olfactory systems, depending on the species. For example the rabbit mammary pheromone is detected by the pups’ main olfactory system. There is a significant overlap even in the size of molecule that is detected by each of the two systems: In the house mouse, while protein pheromones such as darcin are likely only detected by the vomeronasal olfactory system, small molecule pheromones can be detected by either system. The two olfactory systems also have different families of receptors: A mouse pheromone with primer effects on female estrus, 2-heptanone, which is detected by both systems, must thus interact with different receptors in each system.
The main and vomeronasal olfactory systems send their information to different parts of the brain, but these inputs are brought together in the amygdala, a key center for memory and motivation higher up in the brain. So, the two olfactory subsystems may do different things, but there is lots of integration of the inputs—both of pheromone signals and the highly individual smells that allow a mouse to recognize different individuals by smell.
It is highly likely that humans, like other mammals, have pheromones. However, despite the many sites on the Internet offering to sell "pheromones" to make one sexually irresistible, no human pheromones have been chemically identified to date. Sadly there is no evidence for the widely published claims that the molecules androstadienone and estratetraenolare human pheromones of any kind.
There are similar but different problems with the suggestion that women might synchronize their menstrual cycles if living in close quarters, and that this might be mediated by an unidentified pheromone. While some studies have shown the phenomenon of menstrual synchrony, many others have failed to do so. Moreover, the phenomenon has not been found in women in Mali, West Africa, who withdrew together during menstruation, a situation that would have been anticipated to facilitate the effect.
Another misuse of the term pheromone is in so-called pheromone parties, a trend started by an artist in Brooklyn in 2010. At these events, singles sniff numbered T-shirts that have been worn by others at the party to see if they like the smell and photograph themselves with the shirt in a numbered bag to facilitate meeting up later. Although these get-togethers might sound like fun, they are misnamed, because the smells on the shirts are unique to each individual, not species-wide pheromones that trigger the same behavior in every person. The idea behind these parties is based on the surprising observation that mice preferred to mate with mice that were immunologically different from themselves in the major histocompatibility complex (MHC), a difference that can be detected by smell. When a similar experiment was tried with Swiss students sniffing T-shirts, it was discovered that women similarly found the smell of men immunologically different from themselves more attractive. However, subsequent replications of the experiment have not shown the effect consistently and, disappointingly, studies of large-scale human genome data have been equally inconclusive. Clearly, the idea of choosing a potential partner by smell has popular appeal even though the evidence is not there yet.
A human pheromone would need to be a molecule (or combination of molecules) that is characteristic not of a particular individual but of all males or all females, for example, that elicits a reliable behavioral or physiological effect on other individuals. The clearest indication that we may yet find human pheromones in relation to sex is because the smells people give off change after puberty—the sweat and sebaceous glands ramp up their secretions at that point. If we were any other kind of mammal, this change would be a prompt to investigate the smells produced by adults. The techniques for taking and analyzing odor samples are now well established and have been used successfully to identify pheromones in many other mammals. The biggest challenge for work on human pheromones will be the identification of behaviors and physiological responses that will make robust bioassays. Because of our cultural complexity and diversity, influences on human behaviors are notoriously difficult to study. Whatever the future studies on humans, they will need to be guided by the essential steps for identifying a pheromone. There are no shortcuts.
It may be that the first human pheromone to be identified will not have to do with sex but instead with breastfeeding. Finding a mother’s nipple just after birth is a make-or-break moment in the life of a mammal like us, and as I mentioned earlier, other mammals such as rabbits have a pheromone that attracts newborns to the mother’s nipple to suckle.
Developmental ethologist Benoist Schaal and his colleagues from Dijon, France, have found a secretion released from the areola glands around the nipple of human mothers when they produce milk that causes any baby to open its mouth, search for a nipple, and suckle. As yet, we don’t know what molecules might be present—finding and identifying them would be a scientific landmark that would give us more confidence in searching for other possible human pheromones. In addition, if we could synthesize this suspected pheromone, there could be health benefits: If some mothers do not produce enough secretion, supplementing it could perhaps solve problems for nearly half of all newborn babies, who do not feed optimally during their first day, and the many mothers who struggle to breastfeed. The response of the newborn baby to secretions gives a good bioassay, because it is easily testable and infants are also the human life stage least affected by the complexities of learning and culture. I am optimistic that humans may yet join the countless other species of animal that have been shown to use pheromones.
The progress since Butenandt’s identification of the silk moth pheromone molecule in 1959 has been extraordinary. The symbiosis between chemists and biologists continues to reveal pheromones in more species of more kinds of animal than ever before. Apart from the ingenuity of the new chemical techniques for analysis, I am constantly impressed by the ways that pheromones are used to study animal communication: In model animals such as the nematode Caenorhabditis elegans, fruit fly Drosophila melanogaster, many moth species, and the house mouse, researchers can study pheromones at every level—from their production through their perception and neural processing—and from every angle, from genes to development.
Entomologists have long realized that moth pheromones could be used to control pest moths, with the added advantage that natural enemies, like spiders, are left unharmed. In a technique called pheromone mating disruption, synthetic female pheromone is released in crops or orchards in sufficient quantity that male moths cannot find the real females. This technique is being used on more than 10 million hectares around the world. It’s especially effective when all local farmers use it, as the apple growers of Washington State have done to combat the codling moth Cydia pomonella. As is often the case, farmers turned to using pheromones when the moth became resistant to pesticides.
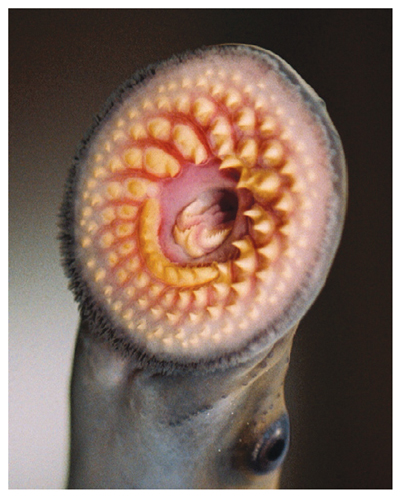
Photograph courtesy of Great Lakes Fishery Commission.
While the use of pheromones has been well established against insect pests, they could also be used to control vertebrate pests and human diseases. For example, invading sea lampreys (Petromyzon marinus), a parasitic jawless fish, have devastated the fisheries of the Great Lakes in North America. Research is ongoing to see if traps baited with the lampreys’ pheromones could replace the toxic chemicals currently being used. Pheromones could also be used to tackle human diseases such as those caused by parasitic nematode worms that use pheromones at crucial stages in their life cycle. One target species is the human parasitic nematode Strongyloides stercoralis, a cause of extensive morbidity in the developing world.

Photograph courtesy of www.epa.gov.
The species specificity of pheromones is one of their big advantages, because using them for pest control has fewer environmental impacts than broad-spectrum pesticides. However, the specificity is a disadvantage for commercial development, because each pest species must be studied individually to identify its pheromones. Unlike the development of broad-spectrum insecticides, the research costs are not spread over many pests. The organizations best placed to work on pheromones are universities and government research laboratories, but these are among those scientific institutions most threatened by spending cuts in many nations. Although it is less profitable for corporations, pheromone-based pest control can be highly cost-effective compared with pesticides and has major benefits to the environment, farmers, and consumers.
Broadcast signals can be "overheard" by unintended recipients. For example, alarm pheromones released by Solenopsis fire ants when fighting ants from other colonies attract unwelcome eavesdroppers: ant-decapitating flies, so named because these parasitic phorid flies lay their eggs in the heads of ants, which eventually fall off. One of these phorid fly species, Pseudacteon tricuspis, might be introduced into the United States as a possible biological control of invasive fire ants.

Photograph by Alexander Wild.
Illegal signalers make counterfeit pheromones as an aggressive form of mimicry. American bolas spider females lure male moths by mimicking the sex pheromones emitted by female moths of that species. When the male moth comes within range, the female Mastophora hutchinsoni spider swings her sticky “bolas” (her web reduced to a ball). If the bolas touches, moths rarely escape.

Photograph courtesy of Kenneth F. Haynes.
Plants can deceive, too. Instead of giving a nectar reward, many orchids dupe male bees into pollinating them by counterfeiting the pheromone of a particular bee species. For example, the European orchid, Ophrys sphegodes, almost perfectly mimics the multicomponent hydrocarbon female sex pheromone blend of the solitary bee, Andrena nigroaenea. Male bees, attracted by the counterfeit pheromone and the visual mimicry of the orchid flower, pick up a pollen pack as they attempt to mate with the flower. They later make the same mistake again and transfer the pollen to another orchid flower of the same species.
Click "American Scientist" to access home page
American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.