Authorship Diplomacy
By Melissa Susan Anderson, Felly Chiteng Kot, Marta A. Shaw, Christine C. Lepkowski, Raymond G. De Vries
Cross-national differences complicate allocation of credit and responsibility
Cross-national differences complicate allocation of credit and responsibility

DOI: 10.1511/2011.90.204
Among scientists, authorship is a very big deal—and for good reason. It not only establishes the record of scientific progress but also stakes a scientist’s claim to originality and priority. As sociologist Robert Merton noted decades ago, recognition for original work is the coin of the realm in science. Authorship is the basis for promotion, tenure, salary, honors and invitations to participate in prestigious initiatives. It is important for collaborating authors to get it right.
Getting it right seems like a simple and straightforward task: Include those who contributed to the project and omit those who did not. Most scientists, however, have encountered situations in which coauthors disagreed about who should be included on a publication or in what order they should be listed. In a recent study, two of us and our colleague Brian Martinson found that 12 percent of midcareer scientists admitted that they had inappropriately assigned authorship credit within the previous three years. Such situations may reflect competitive pressures in science or disputes among authors.
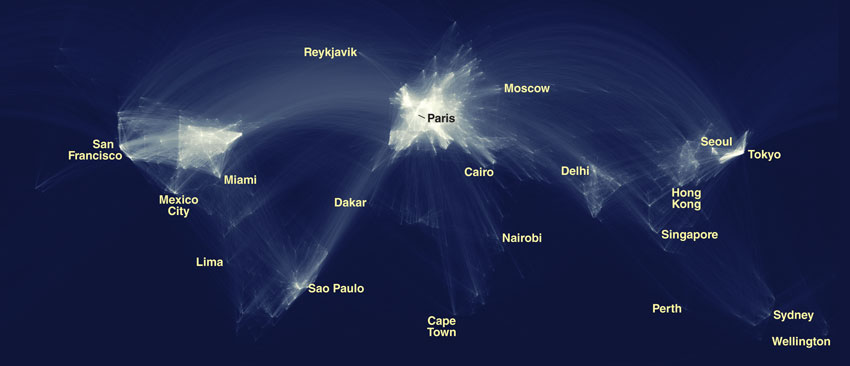
Image courtesy of Olivier Beauchesne, Science-Metrix.
Problems with authorship are complicated enough in domestic research, but they can be particularly thorny in the context of international scientific collaborations. Whether authorship disagreements are more common in international or domestic research is an open question, but some aspects of cross-national collaboration do complicate authorship decisions.
Scientific research is increasingly international in scope and practice. Worldwide, the percentage of science and engineering research articles with authors from more than one country increased from 8 percent in 1988 to 22 percent in 2007, according to the 2010 Science and Engineering Indicators compiled by the U.S. National Science Foundation. Rates of international collaboration as defined in the Indicators are 20 to 30 percent in the United States, China, Japan and India, but around 50 percent in the European Union, in part because recent EU policies and incentives favor international collaboration.
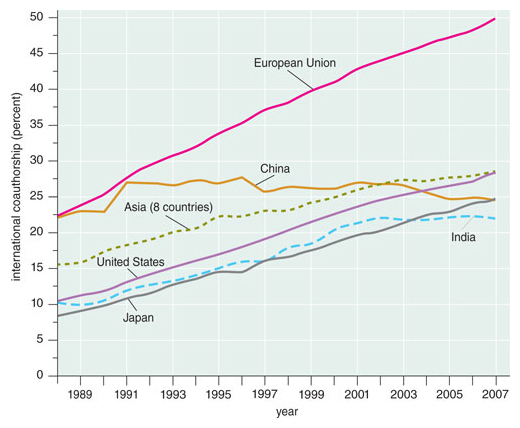
Adapted by Tom Dunne from data published by the U.S. National Science Board.
As we consider authorship issues that arise in these collaborative ventures, we draw on our own and our colleagues’ work in the recently published book International Research Collaborations: Much to be Gained, Many Ways to Get in Trouble and our ongoing research on international scientific collaborations. Specifically, we use material from 10 focus groups and 60 interviews that we conducted over the past year with scientists in the U.S. (and a few outside the U.S.) who are involved in cross-national research collaborations. When we asked these scientists about problematic and beneficial aspects of international research, we inevitably heard about issues with authorship and publication.
One of the most obvious problems in collaborative authorship is omitting authors from a paper. The classic form of omission occurs when two collaborators are in conflict (professional or personal) and one leaves the other’s name off a paper out of spite. Such cases are possible in almost any collaboration—domestic or international. But other forms of omission are more directly linked to cross-national research. Qualifications for authorship, based on scientific contribution or professional status, differ internationally. One scientist told us about working with collaborators in another country who were unwilling to give authorship credit to graduate students simply because of their junior status. He tried to correct the injustice without triggering professional retaliation against the students: “You really almost have to be subversive to help younger people in a way that doesn’t ruin their lives at home, which is not so simple—but I think it is a huge integrity issue.”
We also heard about researchers who left others’ names off publications in order to advance their own careers. In some countries, a senior scientist may feel entitled to take full credit for a junior colleague's work. But the reverse can also happen: Sometimes young scientists train in labs outside their native countries, then publish the results on their own once they return home.
Authors who are omitted without having given their consent often feel wronged, but sometimes authors agree to be left off a publication in exchange for some other form of compensation, usually financial. This arrangement, known as ghost authorship, is a problem in the U.S. as well as in other parts of the world. Last year, Shen Yang of Wuhan University in China released estimates that Chinese academics spent more than $145 million on ghostwritten papers in the previous year. That sum is considerable, especially considering reports of low pay to ghostwriters. For example, Associated Press reporter Gillian Wong last year wrote about a Chinese ghostwriter who received the equivalent of $45 per paper for composing professors’ research articles. One of our focus-group participants commented that pressures and financial rewards for publications increase Chinese academics’ willingness to pay for ghostwriters.
Authors may also remove themselves—either voluntarily or under pressure—from a publication because they fear repercussions for having participated in politically or religiously sensitive research. International collaborators whose research findings may embarrass their governments—for example by exposing weaknesses in health care systems—sometimes ask to be omitted from publications for the sake of their careers.
An omitted author is clearly denied the recognition he or she deserves, but the addition of undeserving authors can also be damaging. Extra names dilute the credit allocated to deserving authors and obscure responsibility for the work. We identify four categories of the added-author problem, distinguishing them according to the motivations for adding an author. These categories overlap to some extent because motivations can be multiple and may not be fully known.
Surprise authorship is when a researcher finds out after publication that his or her name appears on a paper. In some cases, collaborators from different countries do not observe the same practices with regard to coauthorship and review of manuscripts. One scientist told us about a paper published by international colleagues: “I found it by stumbling across the paper in the literature. There it is. This is my name, and there’s the paper, and I have never seen this paper. ”
Gift authorship occurs when someone is given more credit on a paper than he or she deserves. Sometimes a principal investigator decides that it is someone’s turn to be on a publication and arranges for that person's name to appear—even if he or she has not done enough to deserve authorship. In other cases, a senior researcher may decide that his collaborators need publications more than he does, so he allocates publication credit generously (or overly generously) to his collaborators. For instance, in some countries it is common practice to include individuals who have only had administrative oversight. One scientist we interviewed complained about too-liberal inclusion standards among his international colleagues: “Sometimes I am very strict about them, basically saying that I’m not going to allow it. Other times I know there is a political reason why they do it, and so unfortunately I may just let it go by.” Gift authorship is less benign when it involves an expected quid pro quo in the form of future assistance, favors or advantages.
Honorary authorship is often equated with gift authorship, but the motivations are different. Honorary authorship goes to individuals with higher status, as a way of honoring them personally or in their roles as superiors. One scientist told us about working with collaborators in Europe for whom it was standard practice to include “out of respect” the student, the supervisor, the supervisor’s mentor and the department chair.
Often, however, the honor bestowed through unearned authorship is not freely given; rather, it is demanded by supervisors, administrators or funders. This issue came up frequently in our interviews and focus groups. An epidemiologist told us,
People may find that in an international context, if there is a head of the laboratory, that person may expect to go on anything regardless of their contribution or lack of contribution.… So there are issues of sensitivity of where somebody is in terms of the hierarchy.
A focus-group participant explained:
The culture is different. You may be dealing with a researcher who in fact has several layers of bosses. And when it comes to negotiating the dollars, they all get involved. When it comes to publications, they all get to have their name on the paper.
In a 2010 Nature article, author Karen Kaplan relayed suggestions from academics on how to get tenure—including “name a senior department member as a coauthor on your papers if you’re in Europe.”
In legitimizing authorship, a guest author may be listed because of the credibility that his or her position or status will bring to the publication. As one of our respondents put it,
People really want to have your name on their paper, sometimes on papers where you … didn’t even know that they were doing the study. But they’re using a little bit of the reagent you gave them which, you know, you would give anyone freely.… I think that they want your name on a paper because it may legitimize things.
Another biomedical scientist told us about her experience of being named an author of a paper by a research group in another country, to which she had sent plasmids. She thought the study was done incorrectly, and the findings directly contradicted a paper she had published. She continued,
They were upset because what they wanted was my name on the paper so that they could submit it to a journal that was a little bit higher up in the hierarchy, because I already had a reputation in the field. So … this was very uncomfortable because I was saying to them, ‘No, your work isn’t good enough.’ And I was trying to find ways not to say that, but that’s frankly what I felt.
In other cases, legitimizing authorship comes into play when an author is added to mask the illegitimate contributions of others. For example, pharmaceutical firms may recruit researchers to serve as figurehead authors of company-authored papers in order to hide potential conflicts of interest. The figurehead is usually paid well to allow his or her name to appear, frequently replacing the actual authors who may be acknowledged in a footnote or may be absent altogether.
Trouble can also arise as collaborators work out the sequence in which their names will appear on a publication. Disciplinary customs differ, particularly in the significance of the first- and last-named authors. Cross-national teams are often cross-disciplinary as well, and the order of authors can be a point of dispute. One of our focus-group members described collaborating with a high-ranking scientist in another country who insisted on taking the last position to signify, he said, his minimal role on the project. In the journal in question, however, the last position indicated a significant responsibility as the corresponding author for the study. The scientist described it as “a very difficult situation.” Another scientist we interviewed had worked with a team that included a member of a royal family (a princess) who expected to be the first author on every publication in her country, regardless of her contribution.
Some collaborative teams adjust the order of authors to share credit fairly among themselves. They may adopt alphabetical ordering and rotate the alphabetical list in subsequent papers to take turns being first author. Some teams include a note that all authors, or some subset of the authors, contributed equally. Others take advantage of differences in how contributions are measured in different countries. For example, a biological chemist told us,
It worked out nicely.… I was offering them either first or last [author position], thinking that they would of course choose last, but they wanted first, which was perfectly fine with me. That put me last, so in that sense, yeah, it was great.
Plagiarism stakes an implicit authorship claim on someone else’s words or ideas. Recent media attention to plagiarism has alerted scientists to the need to check manuscripts for plagiarized material using increasingly sophisticated software.
Plagiarism among international trainees has long been a concern in many countries. In a 2011 article, research ethicists Elizabeth Heitman of Vanderbilt University and Sergio Litewka of the University of Miami Miller School of Medicine discussed numerous factors that may lead to plagiarism. These include the “normalcy of plagiarism” in some social and cultural environments, vague integrity standards, and rejection of U.S. concepts of originality and intellectual property. Plagiarism may also be a strong temptation for international trainees who have difficulty writing in English but are under pressure to publish in English-language journals. Such problems are not unique to students. In a letter published in Nature in 2007, Turkish physicist Ihsan Yilnaz writes,
For those of us whose mother tongue is not English, using beautiful sentences from other studies on the same subject in our introductions is not unusual.… Borrowing sentences in the part of a paper that simply helps to better introduce the problem should not be seen as plagiarism. Even if our introductions are not entirely original, our results are.
Cultural perspectives on authority also influence writers’ attitudes toward plagiarism. One of our interviewees said, “It took me some years to figure out that there’s an idea at large that anything that comes from authority figures is held in high regard.” She noted that material in the published literature is assumed to come from a person of authority, so students may conclude, “What right do I have to change that person’s words?”
Ready access to articles online has made plagiarism easier. Environmental engineer James Leckie observed in an interview that students in some cultures do not believe they are doing anything wrong when they plagiarize. They argue that “if the authors didn’t want you to use their material, they wouldn’t publish it, but since it’s published and accessible, it should be free for everyone to use in any way.”
A specific issue in the global context is translation plagiarism—that is, translating a publication in whole or in part and publishing the translation as one’s own work without acknowledging the original authors. One U.S. biomedical researcher we interviewed encountered this problem with a collaborator in a developing country. The collaborator asked him to review and endorse a book manuscript, which turned out to be a translation of materials that the U.S. researcher had given to the collaborator, for which the collaborator intended to take sole credit. The U.S. scientist saw that the country in question would derive great value from having the materials in its native language, but he confronted the collaborator about the plagiarism:
I said, ‘I’ve got a real problem with this, because I recognize why you’ve done it and all of this, but the fact is that you have put your name on other people’s words, and what you did is you translated it.’ And he said, ‘Well yes!… What’s the problem with that? And aren’t I your friend? And look at what I have done for you!’ and things like that. It was pretty awkward.
Authorship sits at the intersection between collective effort and individual ambition. Scientists participate in international collaborations for many reasons, including a belief that collaboration will benefit all involved. But the pursuit of individual recognition cannot be completely eliminated. Collaborators must pay careful attention to authorship in order to share credit and responsibility fairly among all team members.
The most helpful way to deal with authorship issues is to agree on general principles for authorship at the beginning of the collaboration and then to agree on authorship of each article when its content is first outlined. A U.S. scientist who is experienced at international collaboration told us, “You have to have the guts to tackle [authorship issues] before you go into it.” Another said,
I found it difficult at first. But it was very clear that if there is any doubt as to how authorship—especially credit—is going to be divvied up, it is better to approach that before, rather than after, just so everybody has a pretty good idea.
It is clear, though, that discussions alone will not clear away all authorship problems. Several resources provide additional guidance. Collaborators should consult the policies of the journals to which they plan to submit their work. International guidance is available through the the Council of Science Editors, the Committee on Publication Ethics and the International Committee on Medical Journal Editors’ Uniform Requirements for Manuscripts Submitted to Biomedical Journals. The Singapore Statement, released in conjunction with the Second World Conference on Research Integrity in 2010, provides a succinct statement that responsible authorship is a duty of researchers worldwide. Scientists can join AuthorAID, a free, international research community that supports researchers from developing countries with services such as networking and mentorship.
The U.S. scientists we interviewed were aware of the ethical complexity of authorship and the importance of meeting high standards in practice. But they were also careful to take their collaborators’ perspectives into account. A focus-group participant said
the problem is that our ignorance of the way these kinds of systems work in other countries can sometimes be really detrimental to the way the research is performed and expressed and published.
Leckie provides a cautionary example, drawn from his own experience working with an Asian collaborator:
I once was designing a research activity with this fellow, and he brought to me very early on several drafts of his part of the proposal. And on it, he had the name of a division head who was associated with the overall program but had nothing to do with his research activity. The division head had no expertise relevant to the proposal and was not going to contribute anything. I told my collaborator, ‘Look, that guy’s not doing anything. Take his name off.’ And he said, ‘Well, I can’t.’ Then I said, ‘Well then, take my name off.’ And so we had a real confrontation, and in the end my collaborator took the name off, and it resulted in an attempt to fire him.
Maintaining the integrity of authorship is complicated in the global context, but the stakes can be high for all concerned. It is worth the time and effort required to get it right.
Click "American Scientist" to access home page
American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.