Bird Song and the Problem of Honest Communication
By William A. Searcy, Stephen Nowicki
How is the honesty of animal signals maintained when exaggeration and bluff are so tempting?
How is the honesty of animal signals maintained when exaggeration and bluff are so tempting?

DOI: 10.1511/2008.70.114
People may at times be tempted to communicate dishonest information to one another: to exaggerate our income to someone we're dating, for example, or, conversely, to understate our income to the Internal Revenue Service. Although morality does play a role in the degree of truth imparted in the messages that we actually deliver, we also use a rational calculation of costs and benefits to decide whether or not to bend the truth. That lie may be worth it to us if the reward outweighs the punishment for getting caught—we are only human, after all.

But when animals communicate, we don't necessarily expect individuals to make decisions based on either moral standards or societal rules. Rather, we expect the rules determining the honesty of a signal—or what researchers studying the topic call signal reliability—to be imposed by natural selection, with costs and benefits ultimately measured in terms of relative reproductive success. How natural selection works to keep animal signals reliable, and the nature of the outcome, is currently a central question in animal behavior.
Signal reliability poses a puzzle in many forms of animal communication, but the nature of this problem is perhaps most easily described by citing an example of aggressive signaling. Suppose two animals are vying for some resource, such as a particularly suitable nesting area, and that they are evenly matched in their fighting ability. Let's also assume that one of these two individuals values the piece of territory more and is therefore willing to fight harder to get it. If the animals resort to fighting, this more aggressive individual will win. Therefore, it seems efficient to settle the contest by signaling: Each individual conveys its true level of aggressiveness—that is, how hard it is willing to fight—and the less-aggressive individual concedes. The outcome is the same as if there had been a fight, but both individuals are spared the often painful price of an actual conflict.
This scenario sounds reasonable enough at first, except that it potentially creates a system that is vulnerable to cheating. If aggressive signals are effective in intimidating rivals, then individuals who exaggerate their aggressiveness will win contests that they would otherwise lose. Cheaters will benefit from their deception, cheating will be favored and spread in the population, and the signal will soon cease to correlate reliably with aggressiveness. Once the signal no longer conveys dependable information, receivers should evolve to ignore it, and once that happens, signalers should cease to give it. In other words, unless there is some way cheating can be held in check, the signaling system is not likely to be maintained by evolution.
The same reasoning applies to signals used in mate attraction. If males honestly advertise their quality, then females will benefit from taking heed of these signals when choosing a mate. If, however, male advertisement has an effect on females' choice of mates, then it would logically follow that selection should act on males to overstate their quality in order to obtain more matings. If exaggeration becomes common enough, these signals will no longer accurately portray the quality of the signaler, receivers should evolve to ignore them, and signalers should cease to produce them.
This apparent dilemma frames a set of key questions that must be answered to understand how reliable communication systems—animal or human—must have evolved. Do the individuals receiving these signals of courtship and aggression respond in ways that benefit the individuals sending them? If so, do these messages contain information about the signalers that is dependable enough to also benefit the receiver? And if the content of the messages being conveyed is genuine, what mechanisms act to keep them that way despite the tempting benefits of dishonesty?
Bird song has been a model for studying the evolution of communication for over half a century. In temperate songbirds, males are the predominant vocalists, singing for the most part in the breeding season, and their songs are thought to play a role both in mate attraction and territory defense. Research indicates these birds' melodies help to attract females and stimulate them to court, copulate and reproduce, and also to help repel rival males from intruding on their territory. In this sense, song is what is known as a "dual function" signal, one that makes it possible for researchers such as us to ask questions about its reliability as a signal of both courtship and aggression. In order to understand what keeps animals honest as they pursue mates or defend their territory, we and our colleagues have been investigating signaling in a particular species of songbird, the song sparrow (Melospiza melodia).
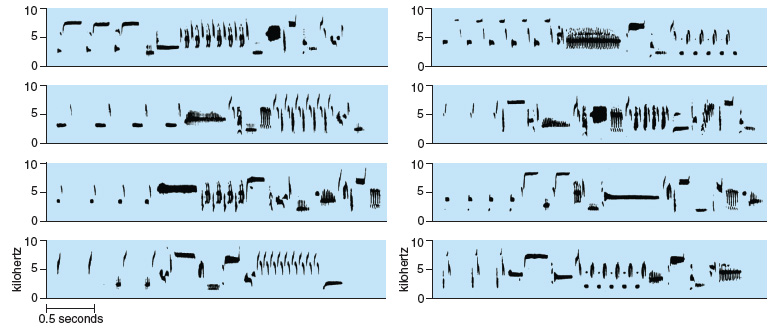
Male song sparrows each possess a repertoire of different versions of their species' songs. For each particular species of songbird, the quality of the acoustic elements, or "notes," that make up a song and the pattern in which these notes are delivered are unique. In many species, however, individuals sing more than one type of song and males vary in how many song types they sing. Repertoire size in male song sparrows ranges from about 5 to 15 types of song, and once established at one year of age the number of song types a male can sing remains set for the rest of his life. Interestingly, the song types that a single male can produce can be so dissimilar that they might seem to have come from different species rather than from the same individual. Nevertheless, the quality of the notes in the song and the pattern in which they are delivered are similar enough within and between repertoires that any song type produced by any song sparrow can be recognized, both by humans and by other song sparrows, as having been produced by that particular species.
Barbara Aulicino.
As variable as the different songs a male sings are, there is no evidence that different song types are specialized for different contexts (for example, interacting with a female versus interacting with a male) or to convey different messages (for example, indicating aggression versus submission). Song sparrows thus fall into the category of songbirds with song types that are interchangeable and in that sense redundant.
Barbara Aulicino
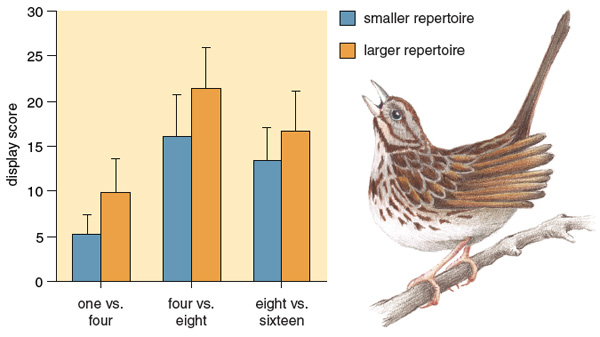
So what do female songbirds listen for in a potential mate? One of us (Searcy), together with Peter Marler, now at the University of California, Davis, found that female song sparrows appear to prefer males with larger song repertoires. In our study, we first treated captive female song sparrows with the sex hormone estradiol to put them in the mood and then played them recordings of song sparrow song. As is true for many species of songbirds, female song sparrows usually respond to a male's song with a solicitation display that typically precedes copulation in nature. In our experiments, we tested female song sparrows for their response to bouts of song sparrow songs composed of differing numbers of song types. First, females were exposed to 32 songs of either four types or one type. Females displayed significantly more for four-type bouts than for single-type bouts. Likewise, later experiments showed that female song sparrows display more in response to 32 songs of eight song types than to 32 songs of four types, and more in response to 64 songs of 16 types than to 64 songs of eight types. So in each case females responded more strongly to larger collections of song types.
As is true for any type of research, conclusions drawn in the laboratory do not always hold true when experiments are taken into the field. Thus, a behavioral preference for large repertoires shown in our initial study does not necessarily mean that repertoire size affects female choice of mates in nature. Indeed, our first attempt to find a relationship between the number of song types a male could sing and mating success outside the laboratory was a failure: Looking across all males in our New York study population, we found no evidence that males with large repertoires paired earlier in the breeding season than did males with small repertoires.
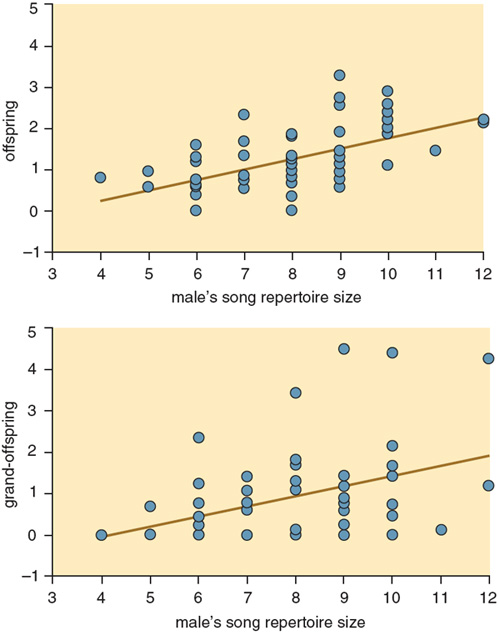
Recently, however, Jane M. Reid of the University of Cambridge and her colleagues did find evidence of the expected relation through a detailed analysis of another population. They examined the song sparrows of Mandarte Island, a small island off Victoria, British Columbia. Because almost all song sparrows born on Mandarte are banded as nestlings, Reid's team of researchers was able to identify and focus on those males that were holding a territory for the first time in a given year. Eliminating older males from the analysis is important, because older males may simply pair again with their female from the previous year, such that their mating success is determined more by history than by their current attributes. When she looked at the first-time breeders, Reid found that the probability that a male would attract a female did in fact increase with the size of his song repertoire.
Together, these field and laboratory results make a strong case that females choose to mate with males who can sing many different types of songs. But these findings raise the question of what female song sparrows gain from such a preference, bearing in mind that it should only evolve if it increases the fitness of the female or her offspring. The best evidence on this issue again comes from Reid and the long-term study of the Mandarte Island population. Along with Peter Arcese and other colleagues from the University of British Columbia, she discovered that the more songs a male can sing, the more offspring, and in turn grand-offspring, he produces over his lifetime. These effects come about in part because males with superior vocal repertoires live longer and thus have more attempts to breed, and in part because they succeed in rearing a greater proportion of their chicks with each breeding attempt. In addition, sons of males with large repertoires themselves give rise to more grand-offspring than do sons of males with small repertoires. These results suggest that females who mate with males with large repertoires gain two types of benefits: The first is a direct benefit to themselves because these males are more successful in rearing young, and the second is an indirect benefit to their young, because these males pass their high fitness to at least their male offspring.
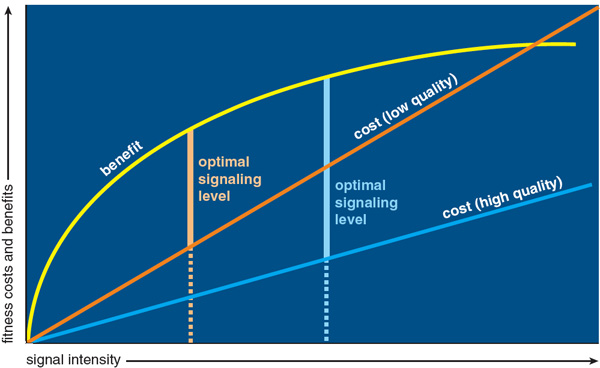
Given that females do indeed base their choice of mate on the number of songs he can sing, what is to keep males from exaggerating this talent in order to appear more attractive? The answer lies in part in Amotz Zahavi's "handicap principle," which states that individuals are less likely to bluff or deceive if the signals they give come at some cost. Game theory (the study of the choice of optimal behavior when the costs and benefits of each option depend on the choices of other individuals) has shown that the handicap principle can work if the cost of the signal increases with the intensity of the signal, and if the increase is more rapid for individuals of low quality than for individuals of high quality. The optimal level of signaling would be higher for individuals of high quality, and if all individuals signal at their optimal level, then the intensity of the signal will reliably reveal the quality of the signaler. One could imagine, for example, that a particular form of signaling might have a substantial energy cost. Individuals who are physically fit and have a lot of energy could then afford to produce the signal at higher rates than could individuals who are less fit and have less energy, and as a result the rate of the signal would reveal the underlying "condition" of the signaler.
Barbara Aulicino
Song repertoire size is unlikely to have an energy cost, however. Singing in general does not seem to use up a lot of energy, and there is no reason to think that singing two songs of different types would be more exhausting than singing two songs of one type. We have suggested that the main costs of song attributes such as repertoire size are developmental: the neural machinery necessary to support song is expensive to manufacture, and therefore only males with superior genetics that have enjoyed good developmental conditions can afford to invest the necessary resources into developing large repertoires of high-quality songs. In support of this notion, we and others have shown that various experimental stresses can negatively affect the development of the brain regions that underlie song learning and production, and hence the structure and complexity of the resulting song.
Barbara Aulicino
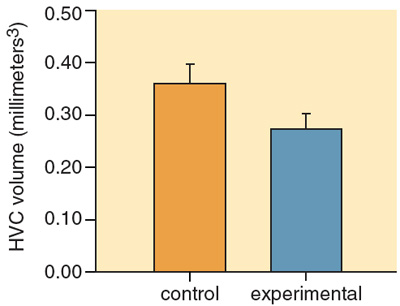
Fernando Nottebohm and his colleagues at of Rockefeller University were the first to identify the specific brain regions, or "nuclei," that control bird song, and since then dozens of researchers have refined and expanded their description. Particularly important is the so-called "higher vocal center" (HVC), an area of the forebrain. The HVC lies at the intersection of two anatomical networks: the descending motor pathway, important to song production, and the anterior forebrain pathway, important to song learning. A number of studies have shown the size of the HVC to be positively correlated with size of the song repertoire, both within and between species. Recently, Ian F. MacDonald and colleagues at the University of Western Ontario manipulated early nutrition in song sparrows and examined effects on HVC size. Subjects were taken from the nest three days after hatching and reared by hand in the laboratory. Birds in the control group were given unlimited food, whereas birds in the experimental group were limited to two-thirds of what the controls ate. When the brains of the subjects were examined at three to four weeks of age, the size of the HVC both on its own and as a proportion of the entire forebrain was greater in the birds given an unlimited diet than in those on a restricted diet.
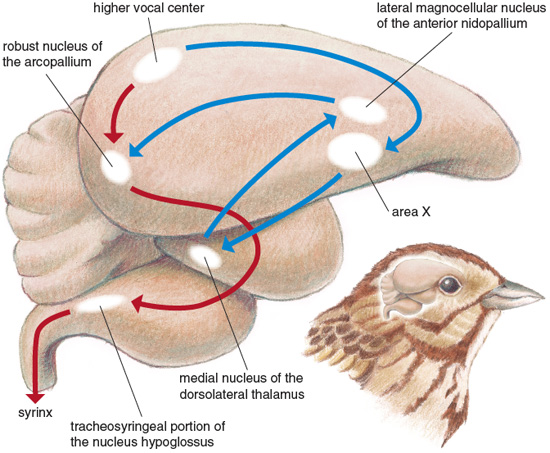
In a separate study, we showed similar effects of early nutrition on HVC size in a close relative of song sparrows, the swamp sparrow (Melospiza georgiana). Both studies thus show that early stress affects the development of a brain region known to be important for the learning and production of bird song.
Barbara Aulicino
Studies in other species of songbirds have taken this research one step further and shown that stress early in life can affect the actual production of adult song. Karen A. Spencer at the University of Bristol and her colleagues have demonstrated that male European starlings (Sturnus vulgaris) given an unpredictable food supply early in life develop reduced repertoire sizes relative to controls. We have shown that swamp sparrows given reduced nutrition before they leave the nest go on to produce songs that are poorer copies of the adult songs they have heard, compared to control males given adequate nutrition. Because these developmental stresses affect not just song but also other aspects of an animal's physique, song is an honest indicator of a male's developmental history and thus of his physical characteristics.
Song can also be an honest indicator of the quality of a male's underlying genetic makeup. Inbreeding provides a convincing measure of this quality, as many studies have shown that individual fitness declines with increased inbreeding. Reid and her colleagues have measured inbreeding in the Mandarte Island population of song sparrows, using pedigrees accumulated over many years of study. They found that the size of song repertoire increases as the level of inbreeding decreases. The idea is that males that are less inbred and are thus genetically superior are as a result buffered from the stresses that influence brain development early in life and its consequent effects on song.
Song sparrows also use song in aggressive signaling between males, and here they employ a number of signaling behaviors that are thought to contain information on aggressiveness. One example is a phenomenon called song type switching. To display a song repertoire a singer must switch among its song types, and varying the frequency with which such switches are made potentially arms the singer with a signal that is not available to species possessing only a single song type. Thus, a songbird could signal a high level of aggressiveness either by increasing or decreasing the frequency of switching; in fact, there is evidence supporting both alternatives in different species. Researchers call song type switching a "conventional" signal, because its meaning is determined by convention rather than by any inherent link between signal and meaning. In song sparrows, a high rate of switching seems, at least superficially, to be associated with aggression. Howard Kramer and Robert Lemon of McGill University showed, for example, that male song sparrows tend to switch more often before and after fights than during undisturbed singing.
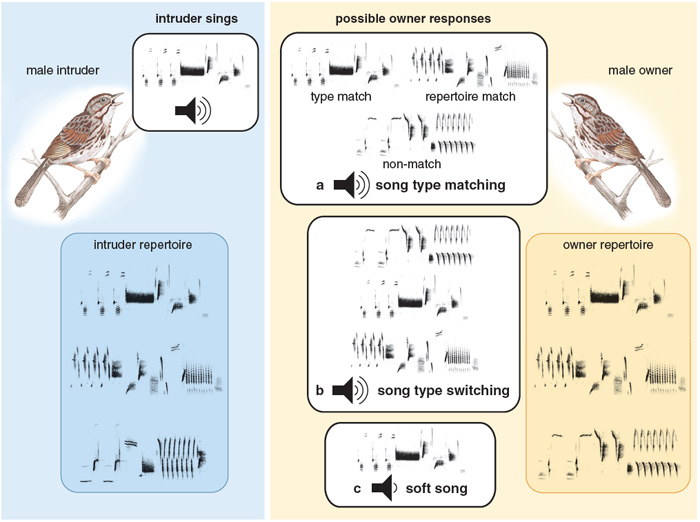
A second way that a male may signal aggression to another male is through a behavior called song type matching. Here one male replies to another with the same song type that the second male has just sung. Matching would be expected to occur occasionally just by chance, but Philip K. Stoddard, Michael D. Beecher and colleagues at the University of Washington showed that male song sparrows matchthe songs that are played from a loudspeaker simulating the intrusion of another bird on their territory at much greater than chance levels. They also discovered a related signaling behavior that represents an even greater degree of sophistication: A male song sparrow may choose to reply to a neighbor not with the song type that the neighbor has just sung, but with another song type that the two males share in both their repertoires. Evidence suggests that "repertoire matching" in this fashion is an intermediate signal, meaning it is less intense than a direct song type match, but more intense than singing a non-matching song type.
Barbara Aulicino
A third, rather puzzling singing behavior that song sparrows perform in aggressive situations is to sing their songs at an especially low volume. For such "soft songs," the amplitude (that is, the strength of the acoustic signal) may be as low as 50 dB SPL ("dB SPL" refers to the sound pressure level in decibels relative to the threshold of human hearing; 50 dB SPL is roughly equivalent to the humming of a refrigerator), compared to 80 to 85 decibels (roughly equivalent to the racket of a blender) for normal song. Investigators have noted soft song in other songbirds in both aggressive and courtship contexts; Margaret Morse Nice, however, in her pioneering monograph on song sparrow behavior, observed soft song only in aggressive situations. The fact that a signal, such as soft song, is most often produced during aggression could indicate that the signal is itself aggressive, in the sense of being more threatening; on the other hand, such evidence could also indicate a submissive meaning, as submissive signals are also more common during aggression than at other times.
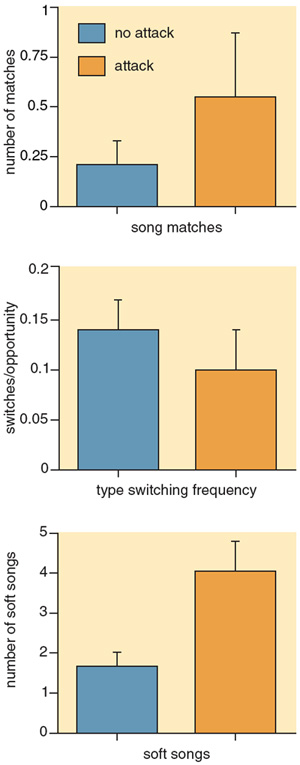
Do any of these singing behaviors convey whether a bird truly intends to attack? To address this question, we devised an experiment in which we first elicited aggressive singing from a male song sparrow by simulating an intrusion on his territory with a previously recorded tape of his song played back for one minute. Next, we recorded the subject's singing for five minutes and then revealed a taxidermic mount of a song sparrow on the bird's territory while playing back another two minutes of his song. The subject was then given 14 minutes to attack or not attack the mount.
Using this study design, we looked for any displays or other behaviors that could be used to predict whether an attack would occur. Clearly, if a display is not reliably associated with a subsequent aggressive act, then it could not be said to act as an aggressive signal. Indeed, we were surprised to find that most of the apparent signaling done by birds in these tests, including type switching and type matching, did not indicate their subsequent behavior. One display, however, did stand out in our analysis—the production of soft song. Males that sang more soft songs were more likely to attack than males that produced few or none.
Given that soft song is indeed the display that most reliably predicts attack in song sparrows, what is to keep males from cheating in order to appear more aggressive? Again, to answer this question one must take another look at the handicap hypothesis, in which the costs of signals are crucial to enforcing their honesty. Scientists have proposed different types of expenses that could be incurred for aggressive signals, but most seem unlikely to apply to soft song. Energy costs, for one, are still unlikely because songs in general, and those of low intensity in particular, take very little energy to produce. The kind of developmental cost discussed earlier for song repertoire size also seems unlikely, because these costs are more likely to apply to signals established over the long term, rather than signals that change over the short term at will. A third possibility is what is called a "vulnerability cost," where the manner in which a signal is produced inherently makes the individual giving it more vulnerable to attack. A vulnerability cost might seem plausible for soft song if, because of its low amplitude, it can only be perceived by a receiver when the singer is close by, making it an unambiguous signal of proximity and thus vulnerability. However, because of the way that sound attenuates with distance, a signal that is soft when it reaches the receiver could be either a low-intensity signal produced near the signaler or a high-intensity signal produced farther away. By contrast, a signal that is loud when it reaches the receiver is actually less ambiguous in conveying that the signaler is close by.
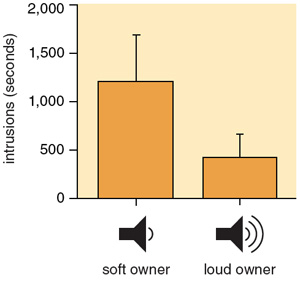
Barbara Aulicino
We have argued that soft song is a reliable aggressive signal because it is an unambiguous and costly signal of attention, not of proximity or vulnerability. Soft song is usually produced quite close to the intended receiver, so that the receiver can both see and hear the singer. The listener then should be able to discern the volume at which the song is produced, and because of its low volume, soft song is unlikely to be audible to any other individuals. Thus by singing at low intensity near a rival, the singer is indicating that its attention is focused solely on that particular bird.
Soft songs are also costly to the singer because they are unlikely to reach other targets. In the case of female receivers, the cost could be a diminished proclivity to mate, as we have shown that female song sparrows find soft song less attractive than songs sung at a normal broadcast amplitude. The more-significant cost of soft song, however, is probably that other male receivers fail to hear a response from the male being challenged, leading neighbors or other potential usurpers to be more likely to intrude on the soft-singer's territory.
A recent study of ours supports the idea that soft song is costly because it limits reception to a single intended receiver. In this experiment, we simulated singing interactions between an intruder and a territory owner, in which the latter sang softly or at a normal level. First we recorded the territory owner and mapped his territory; then we captured and held him temporarily. While the owner was held, we placed two loudspeakers on his territory, and staged a virtual interaction by playing an intruder's song through one speaker and the owner's song from the other. The intruder song was recorded from a male holding a distant territory, and was always played at normal level. In half the trials, the owner's song was also played at normal level, and in the other half the owner's song was played at a level typical of soft song. The result was that intrusions by other male song sparrows were more common and more serious when the simulated owner sang soft songs than when he used loud song. Presumably, other males listening to the interaction from off the territory cannot hear the owner when he uses soft song, and thus cannot tell that he is countering the intruder. Thus by using soft song to signal his focused attention to one intruder, the territory owner sacrifices his ability to ward off other potential intruders.
The fact that animal signals do indeed exist and are maintained in nature implies that they are what researchers call "honest on average." Put simply, even though selection should act on signalers to be deceptive, signals must be honest enough on average that receivers benefit more from paying attention to those signals than from ignoring them. This logic has spurred scientists to investigate what exactly keeps animals from lying. Here, we have suggested two such mechanisms for song sparrows and their signals. In the first case, male song sparrows have difficulty posing as higher quality mates because certain developmental conditions must be met early in life for them to have increased vocal abilities that are attractive to females. In the second case, male song sparrows are unlikely to communicate their intention to attack using soft song if they do not mean business, because by narrowly addressing their signals to one receiver, they abandon their ability to signal to others.
Even though mechanisms such as these exist to ensure that signals remain dependable enough for animals to continue to respond to them, it is still possible for some level of deception to occur. Some song sparrows, for example, might have a larger song repertoire than they "ought" to have given their quality or employ soft songs when they actually are unwilling to attack. Distinguishing cases of deceit from simple error, however, is not a simple matter. Because we cannot know if creatures other than ourselves intend to mislead one another—the question may in fact be meaningless—the criterion that is used to identify dishonesty is whether the signaler benefits from cheating. Thus to demonstrate that deception has occurred, one needs to show both that a signal is in error, and that the signaler benefits from that error. To translate this goal to humans, one would need to prove not only that you lied to say that you are a high-powered hedge-fund manager when you in fact work in the mail room, but also that this deception benefited you by helping you get a date. Searching for falsehoods in this sense may become increasingly important for understanding how communication, including our own, has evolved.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.