Animal Contests as Evolutionary Games
By Mike Mesterton-Gibbons, Eldridge Adams
Paradoxical behavior can be understood in the context of evolutionary stable strategies. The trick is to discover which game the animal is playing
Paradoxical behavior can be understood in the context of evolutionary stable strategies. The trick is to discover which game the animal is playing

DOI: 10.1511/1998.31.334
The science of behavioral ecology thrives on paradoxes, baffling inconsistencies between intuition and evidence that engage our attention and stimulate further investigation. Mathematical models can modify our intuition by showing that apparently sensible explanations are actually problematic, or that seemingly outrageous proposals are downright reasonable. This is especially true in the study of animal contests, where the consequences of an interaction between two animals with opposing interests are difficult to guess.
Consider a famous example known as the "handicap principle." The behavioral ecologist Amotz Zahavi of Tel Aviv University argued that animals with conflicting interests should evolve behavioral displays that are costly to the signaler, even if they lower its chances for survival. By showing that it can endure a handicap, the animal reliably indicates its high quality, a message that other animals do well to respect. For example, on sighting a predator, a gazelle may stott—that is, jump high in the air on all four legs—several times before fleeing, thereby demonstrating that it is in superb physical condition, and that the predator would only waste time and energy by pursuing it. This hypothesis was initially rejected by many partly because it contradicted the biologist's intuition that evolution should favor signals with low costs to the animal producing them, especially since these costly behaviors will be passed onto the signaler's offspring.

Photograph courtesy of Jim Marden, Pennsylvania State University.
Yet formal models of communication revealed that the handicap principle is logically sound under certain conditions. In particular, the magnitude of the handicap must increase with the intensity of the signal, and the cost must be especially damaging for animals of lower quality. For example, if a weak gazelle stotted as vigorously as a strong one, then it would waste what little strength it had and be unable to flee from a pursuing predator. These models have convinced many biologists that a previously unaccepted idea has broad explanatory power.
Efforts to resolve such questions in animal behavior have relied increasingly on collaborations between biologists and mathematicians, using analytical tools called games. In particular, behavioral ecologists use evolutionary games. A game in this context is a mathematical model of strategic interaction, which arises when the outcome of an individual's actions depends on the actions of others.
A game has three components. First, there are at least two interacting individuals, called players. In an evolutionary game played within a single species, the set of players is an ecotype, a population of animals in a given ecological environment. For example, a population of spiders in a grassland habitat and the same species in a riparian habitat form two different ecotypes.
Second, each player has a set of feasible strategies. In an evolutionary game, this set is the same for every player and is constrained by the information structure of the interaction. For example, animals can modify their behavior in different circumstances, such as whether they are owners or intruders, only if they are aware of such roles.
Third, the pattern of interaction must be well defined and accompanied by a formula for how each player's reward from the interaction depends on its strategy and on those of the other players. In evolutionary games, the rewards are measured in terms of expected future reproductive success.
For a game to be useful, it must be possible to identify one or more strategies from among those feasible as the "solution" for a given purpose. In our case, the solution is the behavior that can be expected to evolve by natural selection. If a behavior is fixed in a real population, then it must at least be true that every feasible alternative behavior would yield a lower reward, for otherwise the alternative behavior would have spread into the population. The relevant solution concept, which was introduced a quarter of a century ago by John Maynard Smith of the University of Sussex, is that of an evolutionary stable strategy, a population strategy that yields a higher reward than any feasible mutant strategy (that is, any newly introduced alternative strategy).
Just as a model population is only a caricature of a real population, so a paradox is only a caricature of real ignorance. So, in terms of realism, a game and a paradox are a perfect match. A game theorist strives to unravel the paradox by establishing conditions for an evolutionary stable strategy to exist in the model population, and by analyzing its properties when it does exist. Now, a paradox arises because evidence fails to support intuition, which (assuming the evidence to be sound) can happen only if the intuition relies on a false assumption, albeit an implicit one. So the way to resolve a paradox is to spot the false assumption.
In other words, if a paradox of animal behavior exists, then we have wrongly guessed which game best models how the real population interacts, and to resolve this paradox we must guess again—if necessary, repeatedly—until eventually we guess correctly. Assuming the validity of our solution concept, that is, assuming that the observed behavior corresponds to some evolutionary stable strategy, there are only three things we could be wrong about: the ecotype, the information structure and strategy set or the pattern of interaction and reward. So there are also only three things to be right about. Each can be important, as we illustrate with examples from our recent work on animal contests among damselflies, mantis shrimps and spiders.
A few years ago, Jim Marden of Pennsylvania State University, in collaboration with Jonathan Waage of Brown University, staged a series of territorial contests between male damselflies (Calopteryx maculata). The male pairs in these contests had various, unequal reserves of fat, an indicator of strength. A common expectation for such contests (derived from game-theory models) is that each animal compares its own strength to that of its opponent and withdraws when it judges itself to be the probable loser. This has been confirmed by experimental studies on a variety of animals. The duration of such contests is greatest when the opponents are of nearly equal fighting ability, so that it is more difficult to judge which is likely to win. This is the same reason that many human sporting events are longer and more exciting when teams are evenly matched.
But the duels between the damselflies failed to follow this logic. Although the weaker animal ultimately conceded to its opponent in more than 90 percent of these encounters, Marden found no negative correlation between the duration of a contest and the difference in the relative strengths of the males. Here was a paradox, all the more baffling because the expected relationship had emerged repeatedly in studies of other species. Could there be something amiss in our assumptions about damselfly contests?
One possibly errant assumption is simply the belief that animals can assess one another's strength. Because it is such a good assumption for contests between animals with good visual acuity (such as bighorn sheep), and because so much of a behavioral ecologist's intuition about contest behavior had developed in that context, it was an easy assumption to overlook. But fat is stored internally in insects, so a damselfly cannot directly observe its opponent's reserves. (Marden also found no correlation between fat reserves and traits that surely are observable, such as body length or wing span.) Thus there emerged the intriguing possibility that damselflies do not assess their opponents' strength at all.
Accordingly, in a collaboration with Marden and Lee Dugatkin of the University of Louisville, one of us (Mesterton-Gibbons) developed a model in which a key assumption is that players know only their own reserves. In the resultant game, known as a "war of attrition," a strategy is the proportion of an animal's initial reserves that it is prepared to expend in a prolonged contest over a disputed site. A second key assumption is that the value of winning is proportional to the winner's remaining reserves. The more an animal has left, the more successful it will be in attracting a mate, finding food or defending its territory (and hence in producing surviving offspring). These assumptions determined the strategy set and the reward formula.
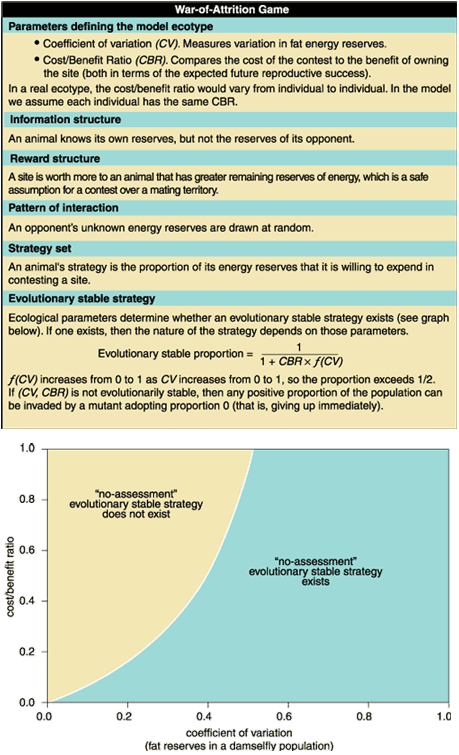
But what about the ecotype? Since a model is merely a caricature of nature, it effectively reduces a real ecological environment to a few parameters, with different values for different ecotypes. In the war-of-attrition game, there are only two such parameters, each a number between zero and one. The first, a coefficient of variation, measures the dispersion of energy reserves about their mean. For example, a coefficient of variation of 0.6 means that one standard deviation in fat reserves is 60 percent of the mean. The second parameter, a cost/benefit ratio, compares the reproductive cost of a spent unit of fat reserves to the eventual winner's reproductive benefit from a saved unit.
With each component of the game identified, we can look for an evolutionary stable strategy. In general, because an evolutionary stable strategy is a population strategy, the ecological parameters determine whether one exists. In this particular game, an evolutionary stable strategy exists (for reasons described below) if the coefficient of variation exceeds a critical threshold, which increases with the cost/benefit ratio but is never much more than about 0.5. The coefficient of variation for Marden's damselflies is 0.51, which would fall below critical if the cost/benefit ratio were between 0.99 and 1. However, in practice the cost/benefit ratio is likely to be considerably less than 0.99. Thus we could assume that fat reserves among damselflies are variable enough that an evolutionary stable strategy involving no assessment of opponents' strength could exist.
At this evolutionary stable strategy, both contestants are prepared to expend at least 50 percent of their initial reserves on the contest, although only the loser actually does so. We can verbally describe the reason for the stability of the no-assessment strategy. Being prepared to expend too small a proportion of initial reserves may mean ceding needlessly to weaker opponents (who would otherwise lose), whereas too high a proportion may mean wasting reserves needlessly against much stronger opponents (who would win in any case). Although animals do not know the reserves of their actual opponents, they achieve a balance between trade-offs by responding to the distribution of reserves among the population.
Marden later staged further damselfly duels in conjunction with Bob Rollins of Pennsylvania State University, and he pooled his data sets. Although the percentage of wins by fatter males fell from 90 percent to 86 percent, it was still remarkably high. Now, judicious approximation is the essence of modeling—effects that are small in a real population are typically absent from a model. For example, we would expect an 86 percent win rate for fatter males in the real world to translate into a 100 percent win rate for fatter males in the model. And this is precisely what happens because both contestants are prepared to deplete their initial reserves by the same proportion. The skinnier one invariably gives up first. In other words, although there is no assessment, the fatter male always wins.
Thus the assessment and no-assessment hypotheses both predict that fatter males always win. But there is also a difference. The assessment hypothesis predicts a negative correlation between strength difference and contest duration. Although, in the pooled data, Marden detected such a correlation in contests exceeding 500 seconds, the variation in strength difference could explain only 14 percent of the variation in contest duration. By contrast, in the no-assessment model, a contest ends when the loser gives up after using a fixed proportion of its reserves. So the no-assessment hypothesis predicts a positive correlation between final loser reserves and contest duration. And, in contests over 500 seconds, the variation in loser reserves was found to explain 29 percent of the variation in duration.
It is tempting to infer from these results that the no-assessment hypothesis is twice as likely to be correct. But we should desist, because 14 percent and 29 percent are both so much less than 100 percent. The results are really inconclusive. Nevertheless, our attempt to resolve the initial paradox has yielded a valuable new insight on animal contest behavior, namely, that a pattern of victory by stronger animals need not imply that strength is being assessed. Moreover, the analysis indicates the kind of population in which we could expect to find such a no-assessment evolutionary stable strategy: one with a high coefficient of variation.
Several years ago, one of us (Adams), in collaboration with Roy Caldwell at the University of California, Berkeley, observed a series of contests between stomatopods, or mantis shrimps, of the species Gonodactylus bredini. These crustaceans inhabit cavities in coral rubble. If one intrudes upon another, the resident often defends its cavity by threatening with a pair of claw-like appendages. These threat displays often deter intruders, so that contests are settled without any physical contact. A surprising observation is that when stomatopods are weakened by molting, so that they are completely unable to fight, they threaten more frequently than animals that are between molts. Furthermore, threats by weaklings often deter much stronger intruders, who would easily win a fight if there were one. In other words, the weaklings bluff. But if the very weakest members of the population can threaten profitably, then why don't all animals threaten? If the display can be given by animals that cannot back it up, then why do their opponents respect it?

Photograph courtesy of Roy L. Caldwell, University of California, Berkeley.
To explore this paradox, we developed a game in which a resident can either threaten or not threaten, and an intruder responds by either attacking or fleeing. We assume that animals vary in weakness, that is, in how much their strength falls short of the maximum strength to be found in the population. We also introduce a reward structure that differs from that assumed by models of "honest" signaling. Specifically, threat displays increase the vulnerability of the signaler to injury inflicted by its opponent. The threat is thus a display of bravado that bears no special cost if the signaler is not attacked, but which adds an additional cost, the threat cost, if it is attacked by a stronger opponent. In addition, if there is a fight, both contestants pay a combat cost, consisting of a fixed cost, which even the strongest animal pays, and a variable cost that increases with weakness at a constant rate of marginal cost.
The threat cost, the fixed cost and the marginal cost together comprise the model ecotype. We also assume that, if there's a fight, the stronger animal wins. Finally, because the molt condition of stomatopods is not externally visible, we assume (as in the war-of-attrition game) that each contestant is unaware of its opponent's strength. So its own strength must determine its behavior.
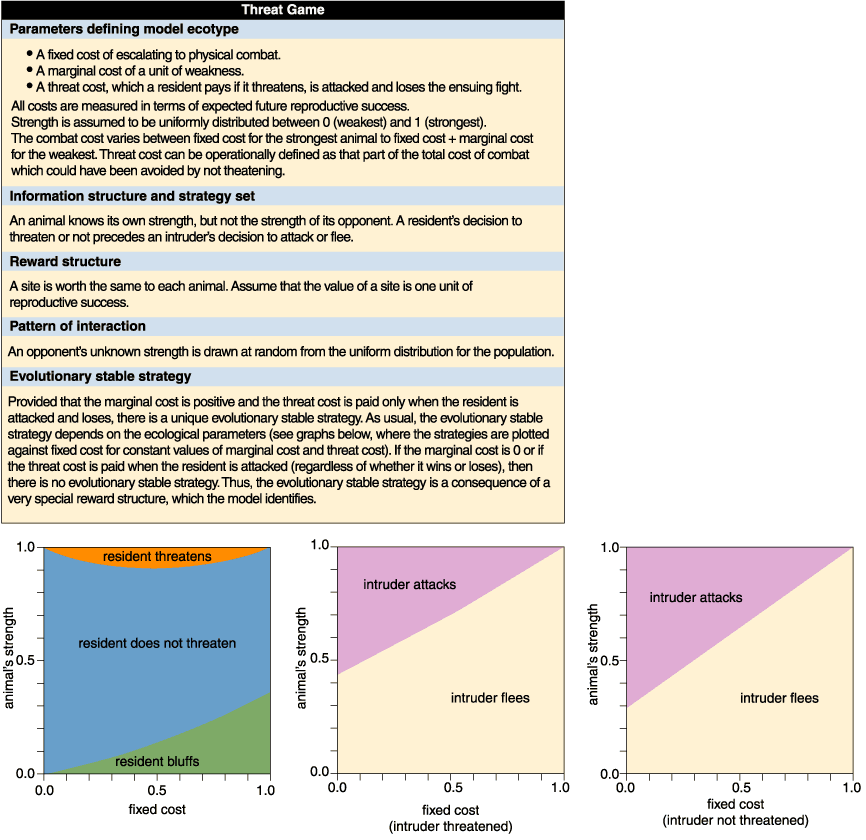
For this particular reward structure, there is always an evolutionary stable strategy at which the weakest and strongest animals both threaten when resident, whereas those of intermediate strength do not (Figure 4, left). Why such a counterintuitive result? All residents face a similar trade-off: Threats deter some opponents without the necessity of fighting, but if the opponent is stronger and chooses to attack, then the threat increases the vulnerability of the signaler. The weaker the signaling animal, the more likely it is to pay the price of its increased vulnerability. That's why animals of intermediate strength cannot afford to threaten, but stronger animals can. This is essentially an instance of the handicap principle. However, the model illustrates a point lacking from previous discussions of handicaps, namely that the benefits of threats are greater for weaker animals, particularly those that have no other means of driving off competitors. These animals would be unlikely to win any of the contests against opponents that are deterred by threats. So the model shows a potential resolution of the paradox: The net costs of bluffing are lower for weak animals than they are for stronger ones. Bluffing therefore forms a part of the evolutionary stable strategy.
As the damselflies exemplify, contests between animals of unequal strength are usually won by the stronger one. Suppose, however, that the animals are equally strong. What advantage could either then have? One possibility in competitions for resources is ownership. Indeed evidence abounds, in species as diverse as baboons, butterflies, lions and red deer, that if animals perceive themselves to be in the separate roles of owner and intruder when contesting a permanent resource, then the owner usually retains it with little serious fighting. There is nothing paradoxical about such respect for ownership. But wouldn't it surprise you if animals avoided serious fighting, not because intruders retreated, but rather because owners fled? That is what appears to happen in a species of Mexican spider, Oecobius civitas.

According to a study by J. Wesley Burgess, populations of this tiny spider live in darkness on the undersides of rocks. If a spider is disturbed from its hiding place, it is apt to enter the hiding place of another, which, if in residence, does not attack the intruder, but instead departs to find a new retreat. A domino effect can ensue, with retreats successively changing ownership until eventually a spider finds one unoccupied. This behavior is inherently paradoxical because, assuming a home-court advantage, the prize always goes to the contestant likelier to lose an actual fight. How can we make sense of this?
As usual, when a behavior is fixed, we expect to understand it as the evolutionary stable strategy of an evolutionary game, with effects that are small in the real population being absent from a model. Thus, we expect butterfly behavior to correspond to an evolutionary stable strategy at which owners always (as opposed to usually) win, and O. civitas behavior to an evolutionary stable strategy at which owners always lose. And, in both cases, we expect fighting to be absent (as opposed to rare). That is, in Maynard Smith's terminology, we expect butterfly behavior to follow a Bourgeois strategy—attack if an owner but be non-aggressive (or display) if an intruder. O. civitas behavior reflects an anti-Bourgeois strategy—display if an owner but attack if an intruder. Likewise, animals that invariably attack, regardless of whether they are owners or intruders, follow a Hawk strategy, and those that display in either role follow a Dove strategy. These four strategies imply an information structure, namely, that animals know their roles.
Using a simple game with these four strategies, Maynard Smith showed that only the Hawk strategy is evolutionarily stable if costs are low enough, whereas the Bourgeois and anti-Bourgeois strategies are evolutionarily stable if costs are high enough. But when two evolutionary stable strategies exist, the population can only be at one of them, and Maynard Smith's game could not say which.
One of us (Mesterton-Gibbons) incorporated Maynard Smith's strategy set into a more elaborate model. Each animal has a finite number of periods in which to find a site, which is essential for reproduction. It does not matter whether the site is acquired early or late, as long as it is owned by the end of the last period. The number of periods is unknown, but all animals have the same probability of surviving predation each period. There are more sites than animals, but the search for a site is random. So a vagrant animal may search in vain, and even if it finds a site it may be occupied. In that case, the animal can either continue to search or contest the site. If it wins, then it becomes an owner, and the previous owner must search again.
There's a catch to this, however. When there's a serious fight, the loser is so seriously injured that it can no longer search, and hence cannot reproduce. Even a winner may suffer injury, and have lower reproductive success than an uninjured animal. So there's a trade-off between the uncertainty of search and the risk of injury. The owner's probability of winning an actual fight is called its resource-holding potential. It is a number between 0.5 and one, indicating that the owner is more likely to win.
Whether animals fight depends on their strategies. A Bourgeois owner and an anti-Bourgeois intruder, or two Hawks, would always fight; an anti-Bourgeois owner and a Bourgeois intruder, or two Doves, would merely display; and a Hawk would always scare off a Dove, in either role. This reward and information structure results in a six-parameter ecotype.
Although the evolutionary stable strategy depends on all six ecological parameters, in terms of our paradox it depends critically on the animal's fighting ability and the probability of its survival. From Figure 6 it is evident that only the Hawk strategy is evolutionarily stable if survival is sufficiently low. Despite the risk of injury, it is better to fight than to hope to find a vacant site before the competition ends. At higher probabilities of survival, however, the Bourgeois or the anti-Bourgeois strategy is evolutionarily stable. To see why, suppose that the resource-holding potential remains constant, but that predation decreases slowly with time (so that the probability of survival increases slowly with time) and that the population tracks its changing environment by continually evolving to a new evolutionary stable strategy. Then we think of the population as a point moving horizontally to the right in Figure 6. As this point migrates, the Hawk strategy must eventually cease to be evolutionarily stable.
Why? Consider a lone Bourgeois mutant in an otherwise Hawk population. Because the Bourgeois animal behaves like a Hawk at home, being an owner has no effect on its reward. So a mutant Bourgeois has a higher reward than the population strategy if a Bourgeois intruder does better than an Hawk intruder. Now, a Bourgeois intruder runs from a Hawk owner, hoping to find an empty site eventually, but a Hawk intruder fights, hoping to win a site immediately. If predation is sufficiently low and the resource-holding potential is sufficiently high, however, the first hope is much more likely to be realized than the second. Thus the Bourgeois fares better than the Hawk and so will spread in the population.
Similarly, a mutant anti-Bourgeois in a Hawk population will behave differently only as an owner. It runs from an intruding Hawk, hoping eventually to find a vacant site, whereas a Hawk owner would fight, hoping to win immediately. If predation and the resource-holding potential are both sufficiently low, then the first of these hopes is much more likely to be realized than the second. Thus the anti-Bourgeois fares better than the Hawk and so will spread in the population.
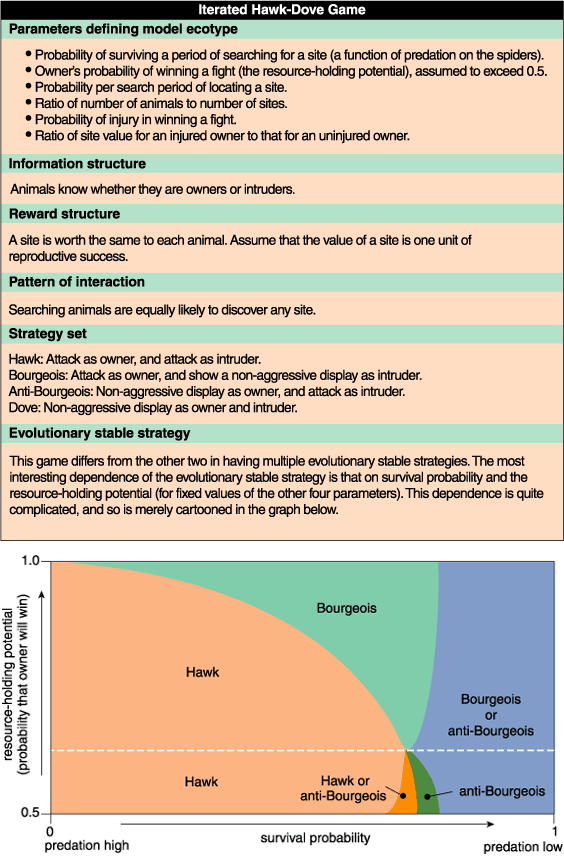
In summary, under decreasing predation, a Hawk population evolves to Bourgeois if the resource-holding potential is relatively large, but the Hawk population evolves to anti-Bourgeois if the resource-holding potential is relatively small (Figure 6). If predation falls sufficiently, then the population will remain at either the Bourgeois or the anti-Bourgeois strategy depending on the initial resource-holding potential. In either case, a non-fighting population evolves from a population of fighters.
To the extent that lower predation increases the chance that not fighting and searching will eventually yield a site without injury, these results agree with those of Maynard Smith. But the new model goes further to suggest a resolution of our paradox. If an ownership asymmetry exists, then the resource-holding potential is almost always high enough to keep the population in a Bourgeois strategy under conditions of low predation. However, there are rare environments in which the resource-holding potential is low enough to keep the population in an anti-Bourgeois strategy under conditions of low predation. Does O. civitas represent such an exception? Perhaps. But without any empirical evidence that the resource-holding potential is indeed low in O. civitas, we can do no more than conjecture.
It is not unusual for an exercise in game theory to remain partially inconclusive. On the one hand, game-theoretic models are valuable because they suggest ways to test new ideas. The damselfly model suggests a test—whether the reserves of the loser correlate positively with contest duration—for an evolutionary stable strategy in which the contestants cannot assess their opponents' strength. The stomatopod model suggests a test—whether animals that threaten and lose pay higher costs than those that lose without threatening—for the idea of an evolutionary stable strategy with partial bluffing. Finally, the spider model suggests a test—whether an intruder is almost as likely as an owner to win an actual fight—for the idea of an anti-Bourgeois evolutionary stable strategy. On the other hand, suggesting a test is not the same thing as conducting it, and the difficulties of doing so should not be underestimated. For example, the only way to measure resource-holding potential is to precipitate actual fights, which may be far from easy in a species as reclusive as the spider O. civitas.
In fact, the difficulties of testing the predictions of evolutionary game theory have led some to question its value. But games are not valuable solely because they suggest ways to test new ideas. They are also valuable because they allow us to explore the logic of a verbal argument rigorously, assuming biologically realistic ecotypes, and to determine when it is true and when it is false. As these cases illustrate, game theory often demonstrates what is difficult to intuit. As a result of exploring damselfly duels, we now understand that victory by stronger animals need not imply that strength is being assessed. As a result of investigating stomatopod strife, we now understand how bluffing can persist at a high frequency. And through analyzing spider spats, we now understand how a non-fighting population of usurpers can evolve, under decreasing predation, from a fighting population that ignores the asymmetry of ownership.
Perhaps non-assessment of strength, high-frequency bluffing and sequential displacement are all remarkably rare in nature. For example, the domino effect has not been observed in colonial spiders other than O. civitas. But it is rarely the commonplace that piques our interest. Rather, it is strange behavior that engages our attention and spurs us on to deeper understanding, even of the commonplace. In that regard, evolutionary games have proved their usefulness over and over again. They have become indispensable analytical tools toward progress in behavioral ecology.
This work was supported by award #9626609 from the National Science Foundation and by a Fellowship in Science and Engineering from the David and Lucile Packard Foundation.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.